

Medaka receptors for somatolactin and growth hormone: phylogenetic paradox among fish growth hormone receptors
- PMID: 16143602
- PMCID: PMC1456111
- DOI: 10.1534/genetics.105.048819
Medaka receptors for somatolactin and growth hormone: phylogenetic paradox among fish growth hormone receptors
Abstract
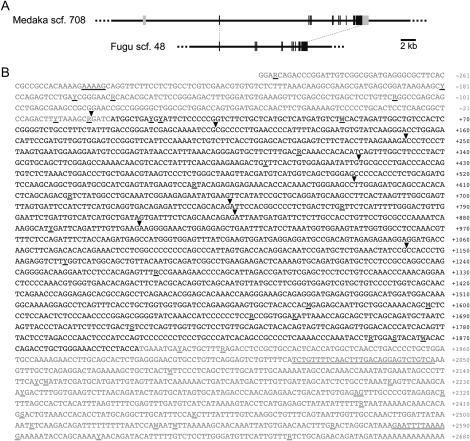
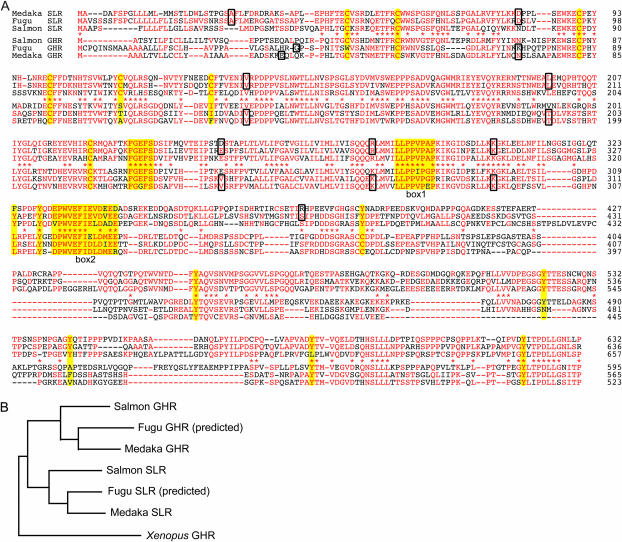
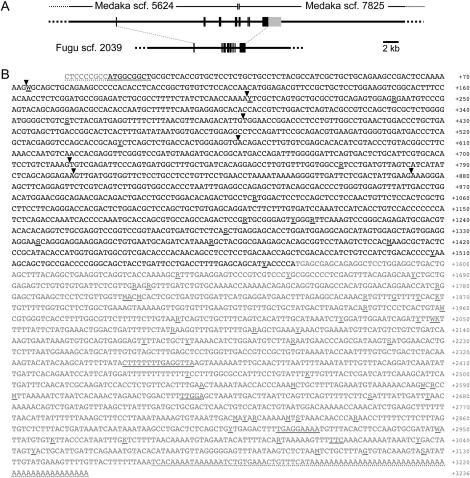
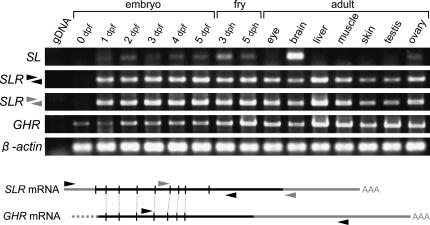
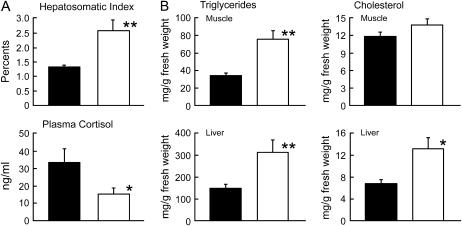
Somatolactin (SL) in fish belongs to the growth hormone/prolactin family. Its ortholog in tetrapods has not been identified and its function(s) remains largely unknown. The SL-deficient mutant of medaka (color interfere, ci) and an SL receptor (SLR) recently identified in salmon provide a fascinating field for investigating SL's function(s) in vivo. Here we isolated a medaka ortholog of the salmon SLR. The mRNA is transcribed in variable organs. Triglycerides and cholesterol contents in the ci are significantly higher than those in the wild type, providing the first evidence of SL's function in suppressing lipid accumulation to organs. Interestingly, phylogenetic comparisons between the medaka SLR and growth hormone receptor (GHR), which is also isolated in this study, in relation to GHRs of other fish, suggested that all GHRs reported from nonsalmonid species are, at least phylogenetically, SLRs. An extra intron inserted in medaka and pufferfish SLRs and flounder and sea bream GHRs also supports their orthologous relationship, but not with tetrapod GHRs. These results may indicate lineage-specific diversification of SLR and GHR functions among fish or just an inappropriate naming of these receptors. Further functional and comparative reassessments are necessary to address this question.
Figures


Similar articles
-
Cloning, phylogenetic analysis and expression of somatolactin and its receptor in Cichlasoma dimerus: their role in long-term background color acclimation.Gen Comp Endocrinol. 2012 Mar 1;176(1):52-61. doi: 10.1016/j.ygcen.2011.12.023. Epub 2011 Dec 29. Gen Comp Endocrinol. 2012. PMID: 22226730
-
Duplication of growth hormone receptor (GHR) in fish genome: gene organization and transcriptional regulation of GHR type I and II in gilthead sea bream (Sparus aurata).Gen Comp Endocrinol. 2005 May 15;142(1-2):193-203. doi: 10.1016/j.ygcen.2004.11.005. Epub 2005 Jan 11. Gen Comp Endocrinol. 2005. PMID: 15862563
-
Cloning of growth hormone, somatolactin, and their receptor mRNAs, their expression in organs, during development, and on salinity stress in the hermaphroditic fish, Kryptolebias marmoratus.Comp Biochem Physiol A Mol Integr Physiol. 2012 Apr;161(4):436-42. doi: 10.1016/j.cbpa.2012.01.004. Epub 2012 Jan 9. Comp Biochem Physiol A Mol Integr Physiol. 2012. PMID: 22261671
-
Somatotropic regulation of fish growth and adiposity: growth hormone (GH) and somatolactin (SL) relationship.Comp Biochem Physiol C Toxicol Pharmacol. 2001 Dec;130(4):435-45. doi: 10.1016/s1532-0456(01)00269-1. Comp Biochem Physiol C Toxicol Pharmacol. 2001. PMID: 11738631 Review.
-
[Growth hormone and prolactin receptors belong to a new receptor family. Biological and medical aspects].Tidsskr Nor Laegeforen. 1993 Feb 28;113(6):725-30. Tidsskr Nor Laegeforen. 1993. PMID: 8465339 Review. Norwegian.
Cited by
-
Growth Hormone Overexpression Disrupts Reproductive Status Through Actions on Leptin.Front Endocrinol (Lausanne). 2018 Mar 27;9:131. doi: 10.3389/fendo.2018.00131. eCollection 2018. Front Endocrinol (Lausanne). 2018. PMID: 29636726 Free PMC article.
-
Evolution of receptors for growth hormone and somatolactin in fish and land vertebrates: lessons from the lungfish and sturgeon orthologues.J Mol Evol. 2007 Oct;65(4):359-72. doi: 10.1007/s00239-007-9035-7. Epub 2007 Oct 5. J Mol Evol. 2007. PMID: 17917757
-
Dual control by a single gene of secondary sexual characters and mating preferences in medaka.BMC Biol. 2009 Sep 29;7:64. doi: 10.1186/1741-7007-7-64. BMC Biol. 2009. PMID: 19788724 Free PMC article.
-
GH, IGF-I and GH receptors mRNA expression in response to growth impairment following a food deprivation period in individually housed cichlid fish Cichlasoma dimerus.Fish Physiol Biochem. 2015 Feb;41(1):51-60. doi: 10.1007/s10695-014-0005-x. Epub 2014 Oct 29. Fish Physiol Biochem. 2015. PMID: 25351458
-
Medaka double mutants for color interfere and leucophore free: characterization of the xanthophore-somatolactin relationship using the leucophore free gene.Dev Genes Evol. 2006 Mar;216(3):152-7. doi: 10.1007/s00427-005-0040-9. Epub 2005 Dec 13. Dev Genes Evol. 2006. PMID: 16344968
References
-
- Amemiya, Y., Y. Sogabe, M. Nozaki, A. Takahashi and H. Kawauchi, 1999. Somatolactin in the white sturgeon and African lungfish and its evolutionary significance. Gen. Comp. Endocrinol. 114: 181–190. - PubMed
-
- Calduch-Giner, J. A., C. Pendon, M. M. Valdivia and J. Perez-Sanchez, 1998. Recombinant somatolactin as a stable and bioactive protein in a cell culture bioassay: development and validation of a sensitive and reproducible radioimmunoassay. J. Endocrinol. 156: 441–447. - PubMed
-
- Calduch-Giner, J. A., H. Duval, F. Chesnel, G. Boeuf, J. Perez-Sanchez et al., 2001. Fish growth hormone receptor: molecular characterization of two membrane-anchored forms. Endocrinology 142: 3269–3273. - PubMed
-
- Calduch-Giner, J. A., M. Mingarro, S. Vega-Rubin de Celis, D. Boujard and J. Perez-Sanchez, 2003. Molecular cloning and characterization of gilthead sea bream (Sparus aurata) growth hormone receptor (GHR). Assessment of alternative splicing. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 136: 1–13. - PubMed
-
- Cavari, B., B. Funkenstein and H. Kawauchi, 2000. Cloning of a second variant of somatolactin-encoding cDNA from the gilthead seabream Sparus aurata. Fish. Physiol. Biochem. 22: 145–150.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
