

The Ferrier Lecture 1995 behind the seen: the functional specialization of the brain in space and time
- PMID: 16147515
- PMCID: PMC1609195
- DOI: 10.1098/rstb.2005.1666
The Ferrier Lecture 1995 behind the seen: the functional specialization of the brain in space and time
Abstract
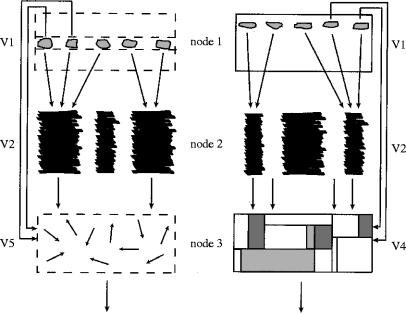
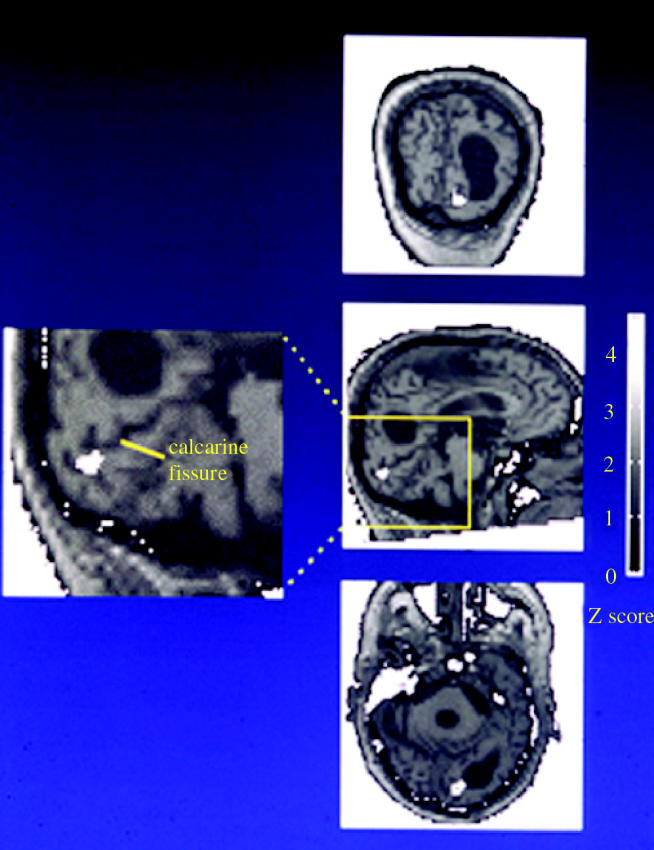
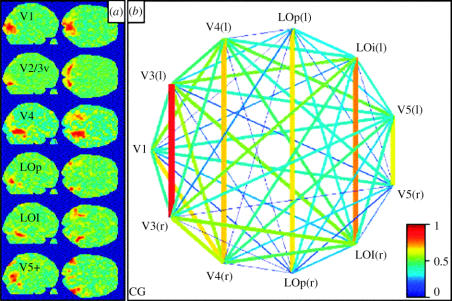
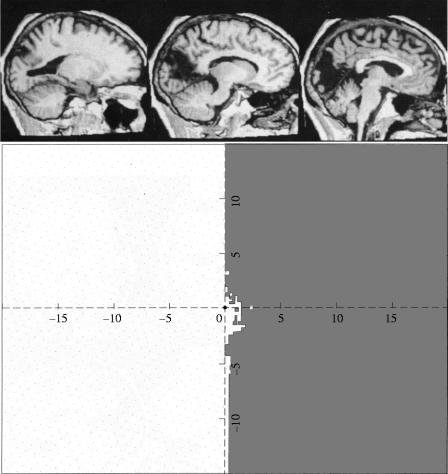
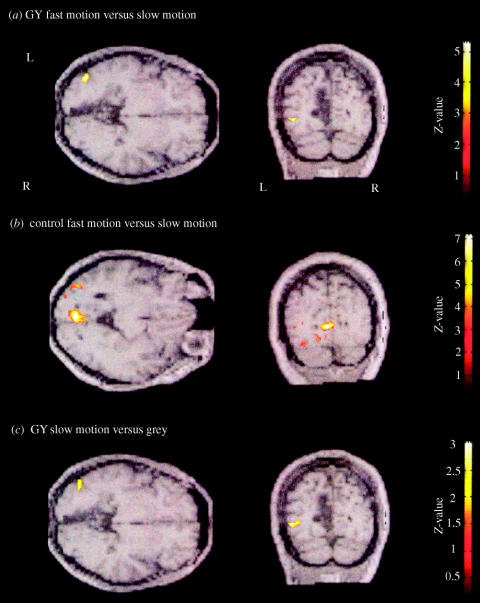
The visual brain consists of many different visual areas, which are functionally specialized to process and perceive different attributes of the visual scene. However, the time taken to process different attributes varies; consequently, we see some attributes before others. It follows that there is a perceptual asynchrony and hierarchy in visual perception. Because perceiving an attribute is tantamount to becoming conscious of it, it follows that we become conscious of different attributes at different times. Visual consciousness is therefore distributed in time. Given that we become conscious of different visual attributes because of activity at different, functionally specialized, areas of the visual brain, it follows that visual consciousness is also distributed in space. Therefore, visual consciousness is not a single unified entity, but consists of many microconsciousnesses.
Figures

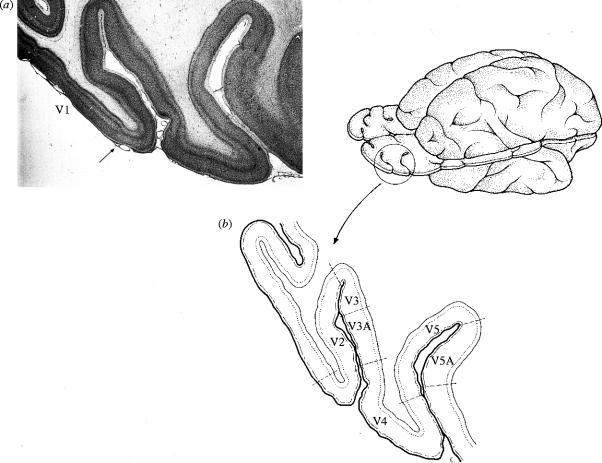
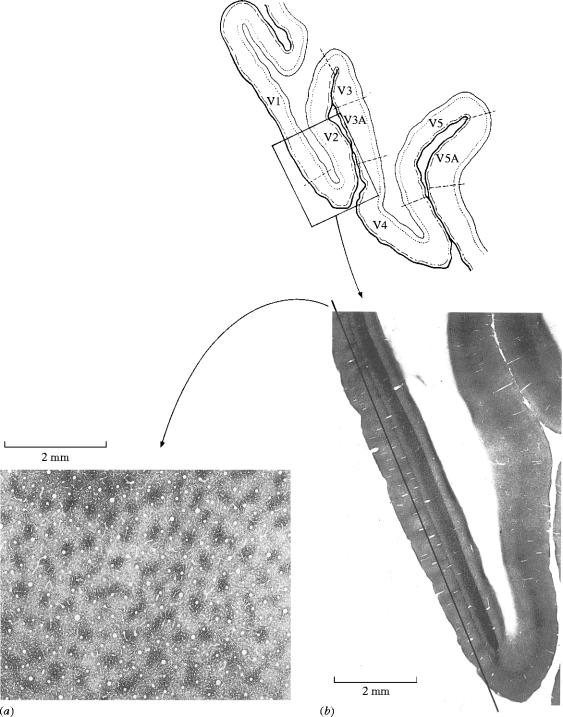
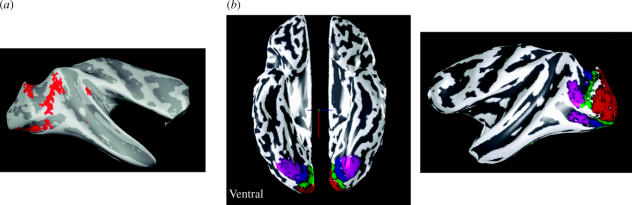

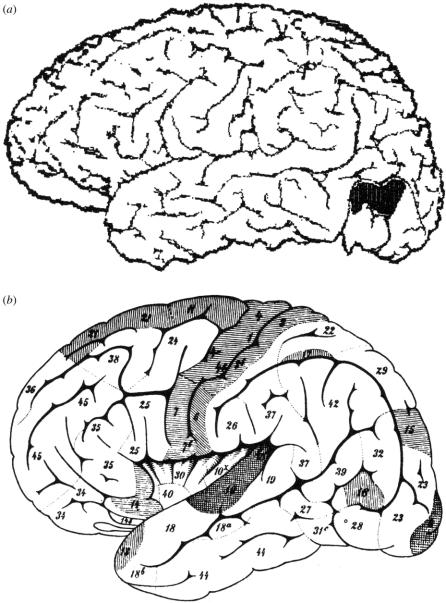
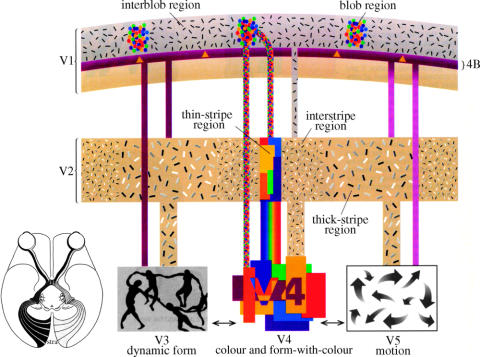


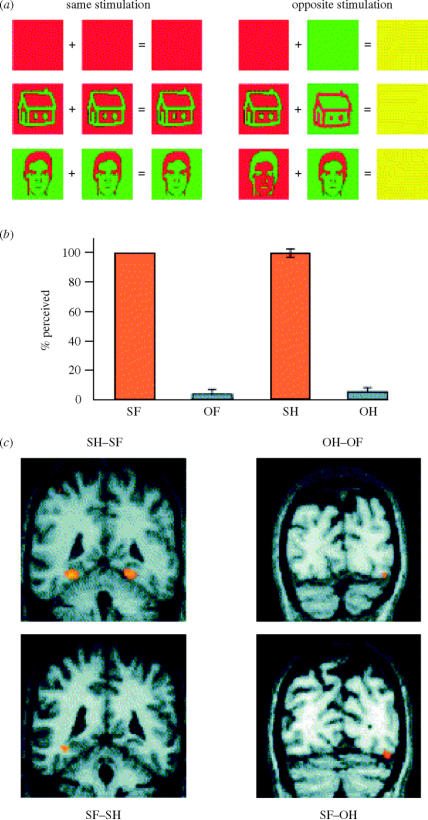
Similar articles
-
Localization and globalization in conscious vision.Annu Rev Neurosci. 2001;24:57-86. doi: 10.1146/annurev.neuro.24.1.57. Annu Rev Neurosci. 2001. PMID: 11283305 Review.
-
The autonomy of the visual systems and the modularity of conscious vision.Philos Trans R Soc Lond B Biol Sci. 1998 Nov 29;353(1377):1911-4. doi: 10.1098/rstb.1998.0343. Philos Trans R Soc Lond B Biol Sci. 1998. PMID: 9854263 Free PMC article. Review.
-
Toward a theory of visual consciousness.Conscious Cogn. 1999 Jun;8(2):225-59. doi: 10.1006/ccog.1999.0390. Conscious Cogn. 1999. PMID: 10448004 Review.
-
The disunity of consciousness.Prog Brain Res. 2008;168:11-8. doi: 10.1016/S0079-6123(07)68002-9. Prog Brain Res. 2008. PMID: 18166382 Review.
-
Vision: modular analysis--or not?Curr Biol. 1999 Feb 11;9(3):R90-2. doi: 10.1016/s0960-9822(99)80057-8. Curr Biol. 1999. PMID: 10021380 Review.
Cited by
-
Cortical responses to consciousness of schematic emotional facial expressions: a high-resolution EEG study.Hum Brain Mapp. 2010 Oct;31(10):1556-69. doi: 10.1002/hbm.20958. Hum Brain Mapp. 2010. PMID: 20143385 Free PMC article.
-
Spatially localized time shifts of the perceptual stream.Front Psychol. 2010 Nov 4;1:181. doi: 10.3389/fpsyg.2010.00181. eCollection 2010. Front Psychol. 2010. PMID: 21833242 Free PMC article.
-
Learning to see words.Annu Rev Psychol. 2012;63:31-53. doi: 10.1146/annurev-psych-120710-100434. Epub 2011 Jul 29. Annu Rev Psychol. 2012. PMID: 21801018 Free PMC article. Review.
-
More than the sum of its parts. Royal Victoria Hospital, Thursday 4th October 2007.Ulster Med J. 2008 Jan;77(1):43-50. Ulster Med J. 2008. PMID: 18269118 Free PMC article. No abstract available.
-
Neural pathways for visual speech perception.Front Neurosci. 2014 Dec 1;8:386. doi: 10.3389/fnins.2014.00386. eCollection 2014. Front Neurosci. 2014. PMID: 25520611 Free PMC article. Review.
References
-
- Albright T.D. Direction and orientation selectivity of neurons in visual area MT of the macaque. J. Neurophysiol. 1984;52:1106–1130. - PubMed
-
- Allman J.M, Kaas J.H. Representation of the visual field in striate and adjoining cortex of the owl monkey (Aotus trivirgatus) Brain Res. 1971;35:89–106. - PubMed
-
- Arnold D.H, Clifford C.W, Wenderoth P. Asynchronous processing in vision: color leads motion. Curr. Biol. 2001;11:596–600. - PubMed
-
- Barbur J.L, Watson J.D, Frackowiak R.S, Zeki S. Conscious visual perception without V1. Brain. 1993;116:1293–1302. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
