

Adhesive mechanisms governing interferon-producing cell recruitment into lymph nodes
- PMID: 16147979
- PMCID: PMC2212867
- DOI: 10.1084/jem.20051035
Adhesive mechanisms governing interferon-producing cell recruitment into lymph nodes
Abstract
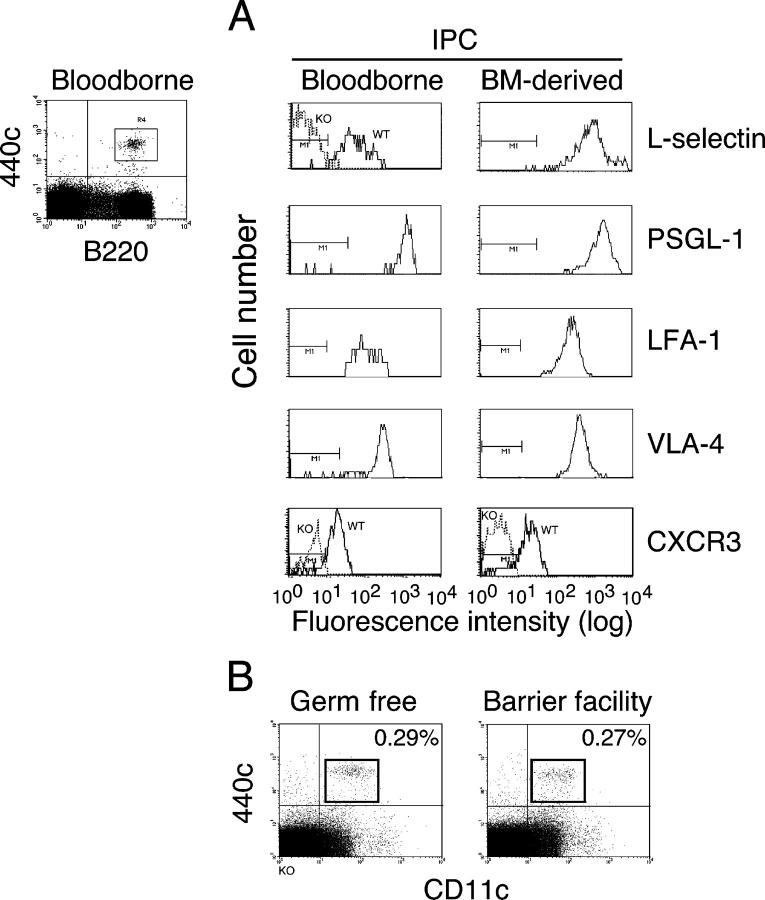
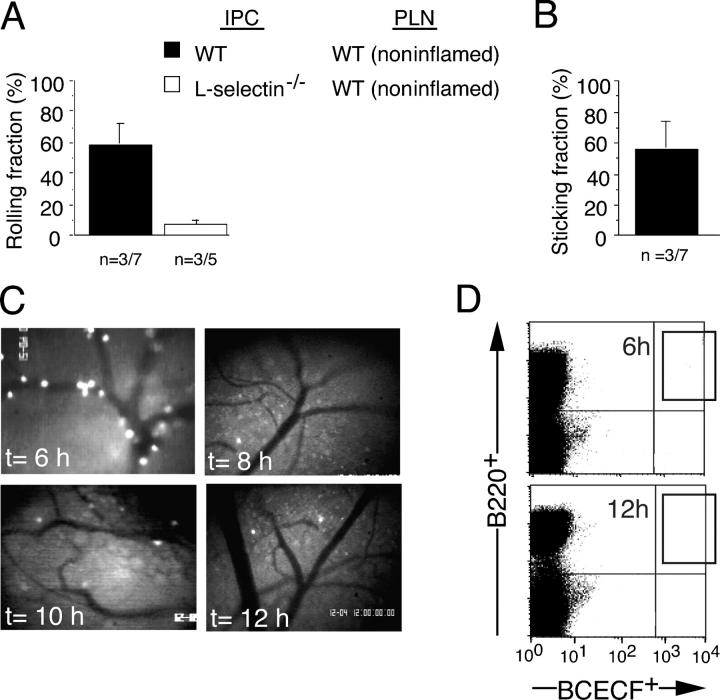
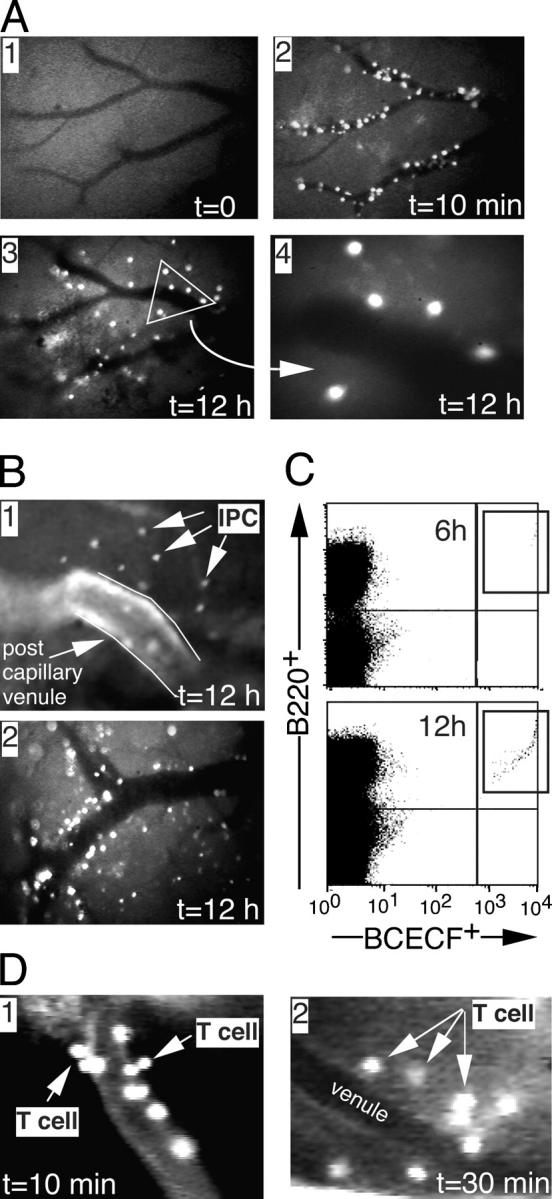
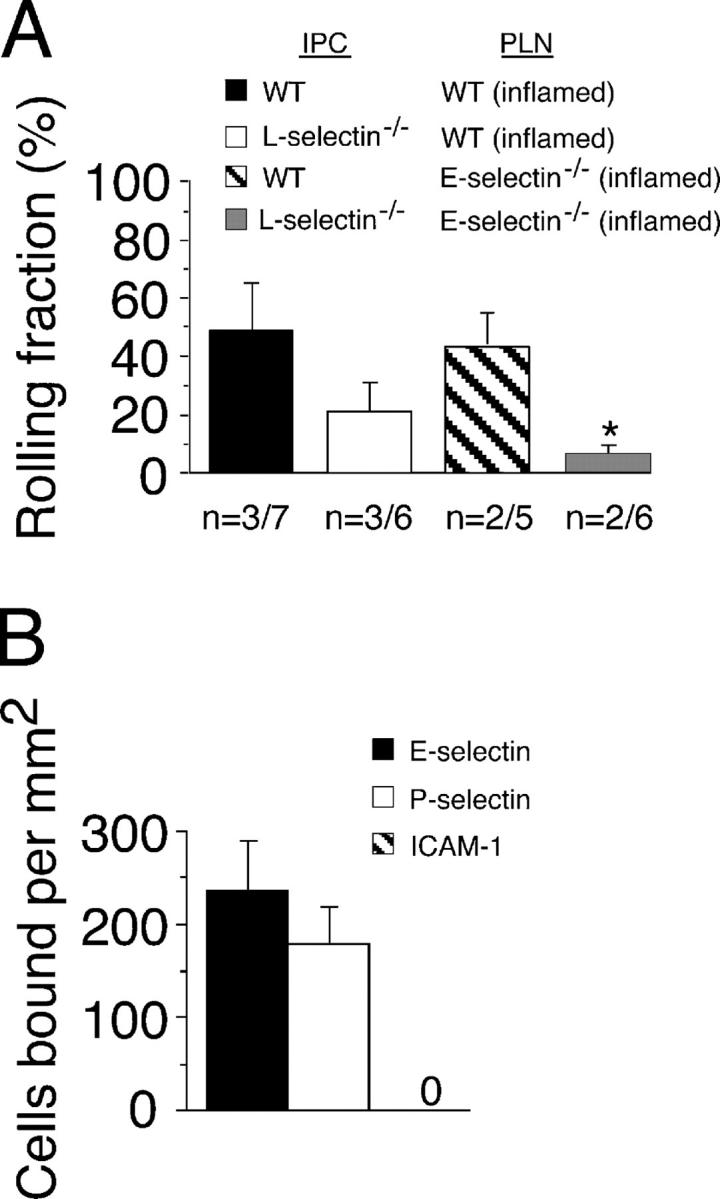
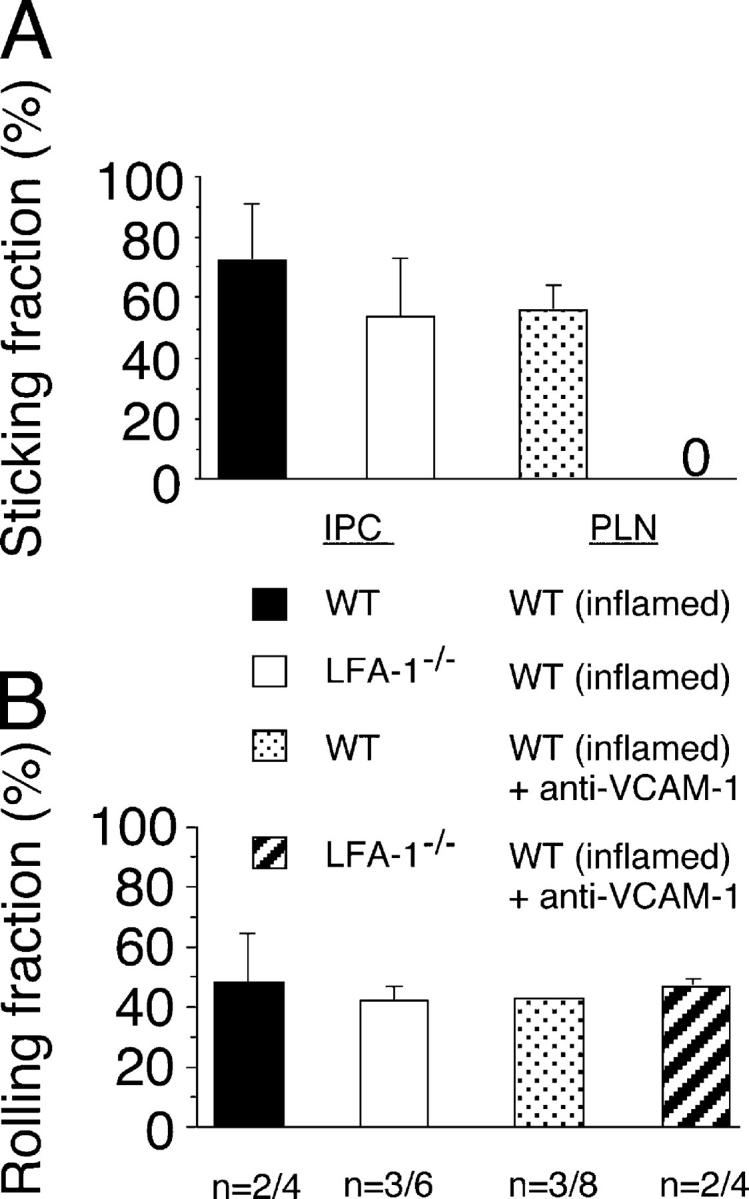
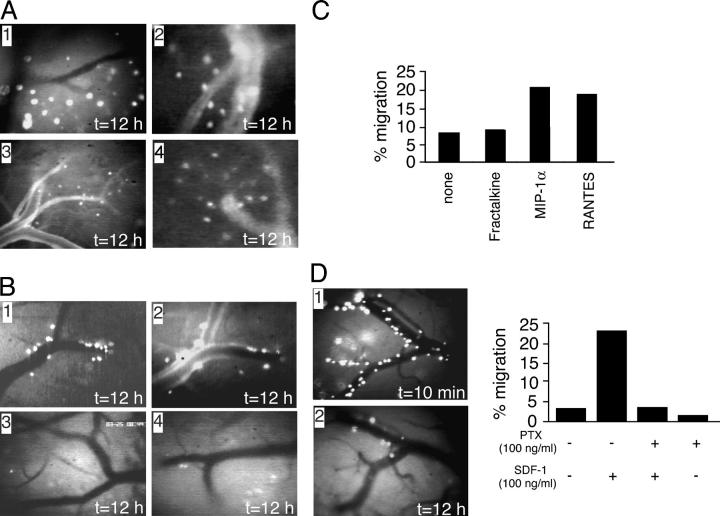
Natural interferon-producing cells (IPCs) are found in peripheral lymph nodes (PLNs), where they support NK cell, T cell, and B cell responses to pathogens. However, their route of entry and the adhesive mechanisms used to gain access to PLNs remain poorly defined. We report that IPCs can enter PLNs via a hematogenous route, which involves a multistep adhesive process, and that transmigration is enhanced by inflammation. Results indicate that L-selectin on IPCs is required for efficient attachment and rolling on high endothelial venules in vivo in both nonstimulated and inflamed PLNs. IPCs, however, also possess functional ligands for E-selectin that contribute to this process only in the latter case. In conjunction with selectin-mediated adhesion, both beta(1)- and beta(2)-integrins participate in IPC attachment to the inflamed vessel wall, whereas chemotaxis relies in part on the chemokine receptor CCR5. Identification of the adhesive machinery required for IPC trafficking into PLNs may provide opportunities to regulate immune responses reliant on the activity of these cells.
Figures
References
-
- Colonna, M., G. Trinchieri, and Y.J. Liu. 2004. Plasmacytoid dendritic cells in immunity. Nat. Immunol. 5:1219–1226. - PubMed
-
- Liu, Y.J. 2005. IPC: professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors. Annu. Rev. Immunol. 23:275–306. - PubMed
-
- Iwasaki, A., and R. Medzhitov. 2004. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 5:987–995. - PubMed
-
- Siegal, F.P., N. Kadowaki, M. Shodell, P.A. Fitzgerald-Bocarsly, K. Shah, S. Ho, S. Antonenko, and Y.J. Liu. 1999. The nature of the principal type 1 interferon-producing cells in human blood. Science. 284:1835–1837. - PubMed
-
- Cella, M., F. Facchetti, A. Lanzavecchia, and M. Colonna. 2000. Plasmacytoid dendritic cells activated by influenza virus and CD40L drive a potent TH1 polarization. Nat. Immunol. 1:305–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
