

Evidence that the Tfg1/Tfg2 dimer interface of TFIIF lies near the active center of the RNA polymerase II initiation complex
- PMID: 16147988
- PMCID: PMC1201334
- DOI: 10.1093/nar/gki825
Evidence that the Tfg1/Tfg2 dimer interface of TFIIF lies near the active center of the RNA polymerase II initiation complex
Abstract
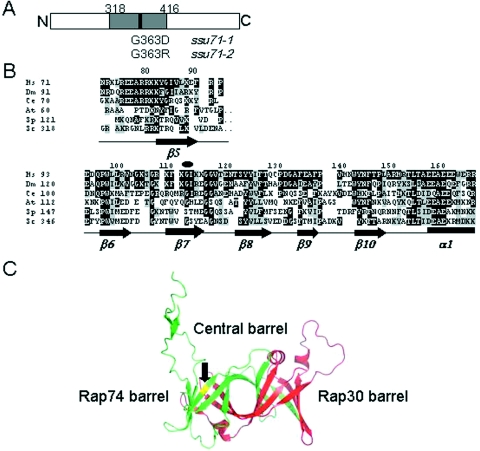
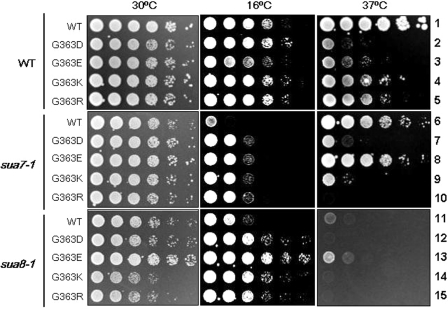
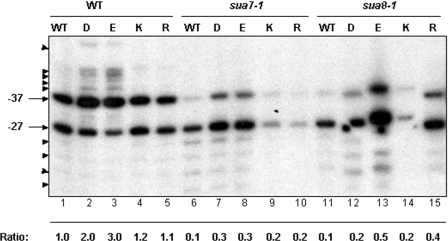
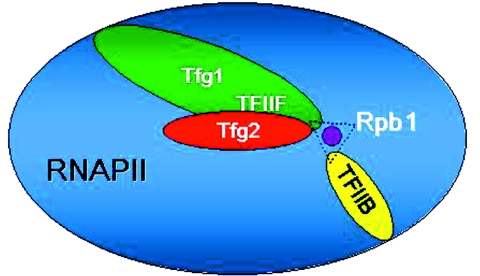
The ssu71 alleles of the TFG1 gene, which encodes the largest subunit of TFIIF, were isolated as suppressors of a TFIIB defect that affects the accuracy of transcription start site selection in the yeast Saccharomyces cerevisiae. Here we report that ssu71-1 also suppresses the cell growth and start site defects associated with an altered form of the Rpb1 subunit of RNA polymerase II (RNAP II). The ssu71-1 and ssu71-2 alleles were cloned and found to encode single amino acid replacements of glycine-363, either glycine to aspartic acid (G363D) or glycine to arginine (G363R). Two other charged replacements, G363E and G363K, were constructed by site-directed mutagenesis and suppress both TFIIB E62K and Rpb1 N445S, whereas neither G363A nor G363P exhibited any effect. G363 is phylogenetically conserved and its counterpart in human TFIIF (RAP74 G112) is located within the RAP74/RAP30 dimerization domain. We propose that the TFIIF dimerization domain is located in proximity to the B-finger of TFIIB near the active center of RNAP II where the TFIIB-TFIIF-RNAP II interface plays a key role in start site selection.
Figures

Similar articles
-
Amino acid substitutions in yeast TFIIF confer upstream shifts in transcription initiation and altered interaction with RNA polymerase II.Mol Cell Biol. 2004 Dec;24(24):10975-85. doi: 10.1128/MCB.24.24.10975-10985.2004. Mol Cell Biol. 2004. PMID: 15572698 Free PMC article.
-
Functions of Saccharomyces cerevisiae TFIIF during transcription start site utilization.Mol Cell Biol. 2008 Jun;28(11):3757-66. doi: 10.1128/MCB.02272-07. Epub 2008 Mar 24. Mol Cell Biol. 2008. PMID: 18362165 Free PMC article.
-
Position of the general transcription factor TFIIF within the RNA polymerase II transcription preinitiation complex.EMBO J. 2010 Feb 17;29(4):706-16. doi: 10.1038/emboj.2009.386. Epub 2009 Dec 24. EMBO J. 2010. PMID: 20033062 Free PMC article.
-
Rethinking the role of TFIIF in transcript initiation by RNA polymerase II.Transcription. 2012 Jul-Aug;3(4):156-9. doi: 10.4161/trns.20725. Epub 2012 Jul 1. Transcription. 2012. PMID: 22771986 Free PMC article. Review.
-
Conservation between the RNA polymerase I, II, and III transcription initiation machineries.Mol Cell. 2012 Feb 24;45(4):439-46. doi: 10.1016/j.molcel.2012.01.023. Mol Cell. 2012. PMID: 22365827 Review.
Cited by
-
Structural visualization of key steps in human transcription initiation.Nature. 2013 Mar 28;495(7442):481-6. doi: 10.1038/nature11991. Epub 2013 Feb 27. Nature. 2013. PMID: 23446344 Free PMC article.
-
The functions of TFIIF during initiation and transcript elongation are differentially affected by phosphorylation by casein kinase 2.J Biol Chem. 2011 Jul 1;286(26):23160-7. doi: 10.1074/jbc.M110.205658. Epub 2011 May 12. J Biol Chem. 2011. PMID: 21566144 Free PMC article.
-
Mechanism of start site selection by RNA polymerase II: interplay between TFIIB and Ssl2/XPB helicase subunit of TFIIH.J Biol Chem. 2012 Jan 2;287(1):557-567. doi: 10.1074/jbc.M111.281576. Epub 2011 Nov 11. J Biol Chem. 2012. PMID: 22081613 Free PMC article.
-
Architecture of the RNA polymerase II-TFIIF complex revealed by cross-linking and mass spectrometry.EMBO J. 2010 Feb 17;29(4):717-26. doi: 10.1038/emboj.2009.401. Epub 2010 Jan 21. EMBO J. 2010. PMID: 20094031 Free PMC article.
-
RNA polymerase II mutations conferring defects in poly(A) site cleavage and termination in Saccharomyces cerevisiae.G3 (Bethesda). 2013 Feb;3(2):167-80. doi: 10.1534/g3.112.004531. Epub 2013 Feb 1. G3 (Bethesda). 2013. PMID: 23390594 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
