

Hyperpolarization-activated cation current (Ih) is an ethanol target in midbrain dopamine neurons of mice
- PMID: 16148268
- PMCID: PMC1454360
- DOI: 10.1152/jn.00682.2005
Hyperpolarization-activated cation current (Ih) is an ethanol target in midbrain dopamine neurons of mice
Abstract
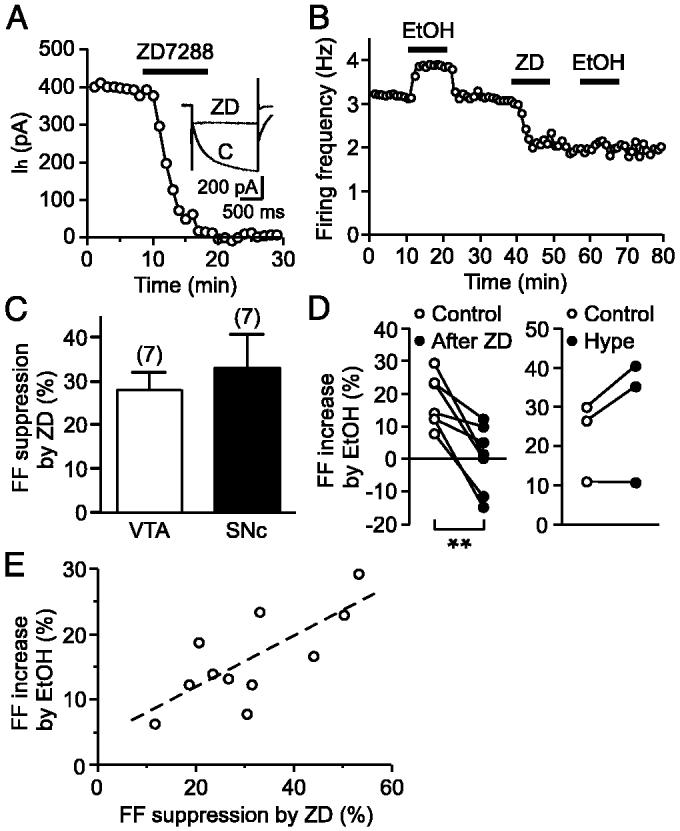
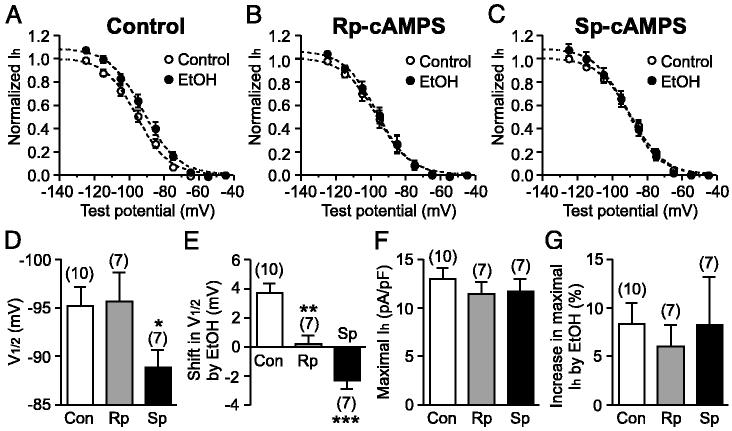
Ethanol stimulates the firing activity of midbrain dopamine (DA) neurons, leading to enhanced dopaminergic transmission in the mesolimbic system. This effect is thought to underlie the behavioral reinforcement of alcohol intake. Ethanol has been shown to directly enhance the intrinsic pacemaker activity of DA neurons, yet the cellular mechanism mediating this excitation remains poorly understood. The hyperpolarization-activated cation current, Ih, is known to contribute to the pacemaker firing of DA neurons. To determine the role of Ih in ethanol excitation of DA neurons, we performed patch-clamp recordings in acutely prepared mouse midbrain slices. Superfusion of ethanol increased the spontaneous firing frequency of DA neurons in a reversible fashion. Treatment with ZD7288, a blocker of Ih, irreversibly depressed basal firing frequency and significantly attenuated the stimulatory effect of ethanol on firing. Furthermore, ethanol reversibly augmented Ih amplitude and accelerated its activation kinetics. This effect of ethanol was accompanied by a shift in the voltage dependence of Ih activation to more depolarized potentials and an increase in the maximum Ih conductance. Cyclic AMP mediated the depolarizing shift in Ih activation but not the increase in the maximum conductance. Finally, repeated ethanol treatment in vivo induced downregulation of Ih density in DA neurons and an accompanying reduction in the magnitude of ethanol stimulation of firing. These results suggest an important role of Ih in the reinforcing actions of ethanol and in the neuroadaptations underlying escalation of alcohol consumption associated with alcoholism.
Figures





Comment in
-
Queer currents, steady rhythms, and drunken da neurons. Focus on "hyperpolarization-activated cation current (Ih) is an ethanol target in midbrain dopamine neurons of mice".J Neurophysiol. 2006 Feb;95(2):585-6. doi: 10.1152/jn.00957.2005. J Neurophysiol. 2006. PMID: 16424449 No abstract available.
References
-
- Ahlenius S, Carlsson A, Engel J, Svensson T, Sodersten P. Antagonism by alpha methyltyrosine of the ethanol-induced stimulation and euphoria in man. Clin Pharmacol Ther. 1973;14:586–591. - PubMed
-
- Appel SB, Liu Z, McElvain MA, Brodie MS. Ethanol excitation of dopaminergic ventral tegmental area neurons is blocked by quinidine. J Pharmacol Exp Ther. 2003;306:437–446. - PubMed
-
- Asher O, Cunningham TD, Yao L, Gordon AS, Diamond I. Ethanol stimulates cAMP-responsive element (CRE)-mediated transcription via CRE-binding protein and cAMP-dependent protein kinase. J Pharmacol Exp Ther. 2002;301:66–70. - PubMed
-
- Brodie MS. Increased ethanol excitation of dopaminergic neurons of the ventral tegmental area after chronic ethanol treatment. Alcohol Clin Exp Res. 2002;26:1024–1030. - PubMed
-
- Brodie MS, Appel SB. The effects of ethanol on dopaminergic neurons of the ventral tegmental area studied with intracellular recording in brain slices. Alcohol Clin Exp Res. 1998;22:236–244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources