

Speciation and gene flow between snails of opposite chirality
- PMID: 16149849
- PMCID: PMC1182688
- DOI: 10.1371/journal.pbio.0030282
Speciation and gene flow between snails of opposite chirality
Abstract
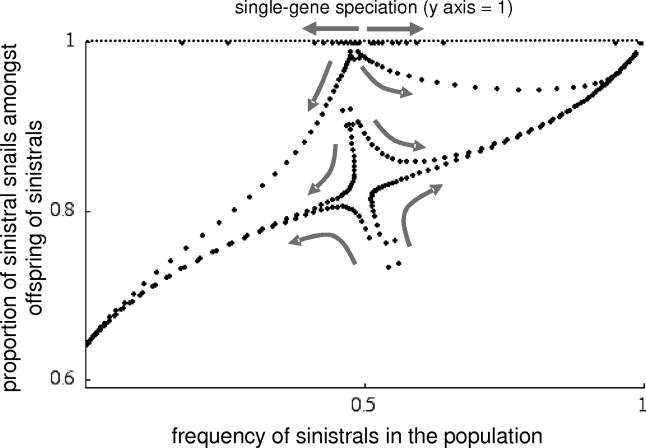
Left-right asymmetry in snails is intriguing because individuals of opposite chirality are either unable to mate or can only mate with difficulty, so could be reproductively isolated from each other. We have therefore investigated chiral evolution in the Japanese land snail genus Euhadra to understand whether changes in chirality have promoted speciation. In particular, we aimed to understand the effect of the maternal inheritance of chirality on reproductive isolation and gene flow. We found that the mitochondrial DNA phylogeny of Euhadra is consistent with a single, relatively ancient evolution of sinistral species and suggests either recent "single-gene speciation" or gene flow between chiral morphs that are unable to mate. To clarify the conditions under which new chiral morphs might evolve and whether single-gene speciation can occur, we developed a mathematical model that is relevant to any maternal-effect gene. The model shows that reproductive character displacement can promote the evolution of new chiral morphs, tending to counteract the positive frequency-dependent selection that would otherwise drive the more common chiral morph to fixation. This therefore suggests a general mechanism as to how chiral variation arises in snails. In populations that contain both chiral morphs, two different situations are then possible. In the first, gene flow is substantial between morphs even without interchiral mating, because of the maternal inheritance of chirality. In the second, reproductive isolation is possible but unstable, and will also lead to gene flow if intrachiral matings occasionally produce offspring with the opposite chirality. Together, the results imply that speciation by chiral reversal is only meaningful in the context of a complex biogeographical process, and so must usually involve other factors. In order to understand the roles of reproductive character displacement and gene flow in the chiral evolution of Euhadra, it will be necessary to investigate populations in which both chiral morphs coexist.
Figures







Similar articles
-
The convoluted evolution of snail chirality.Naturwissenschaften. 2005 Nov;92(11):504-15. doi: 10.1007/s00114-05-0045-2. Epub 2005 Oct 11. Naturwissenschaften. 2005. PMID: 16217668 Review.
-
Single-gene speciation: Mating and gene flow between mirror-image snails.Evol Lett. 2017 Nov 21;1(6):282-291. doi: 10.1002/evl3.31. eCollection 2017 Dec. Evol Lett. 2017. PMID: 30283656 Free PMC article.
-
Chiral speciation in terrestrial pulmonate snails.PLoS One. 2012;7(4):e34005. doi: 10.1371/journal.pone.0034005. Epub 2012 Apr 20. PLoS One. 2012. PMID: 22532825 Free PMC article.
-
The effect of coil phenotypes and genotypes on the fecundity and viability of Partula suturalis and Lymnaea stagnalis: implications for the evolution of sinistral snails.J Evol Biol. 2009 Aug;22(8):1624-35. doi: 10.1111/j.1420-9101.2009.01770.x. Epub 2009 Jun 11. J Evol Biol. 2009. PMID: 19523040
-
Flipping Shells! Unwinding LR Asymmetry in Mirror-Image Molluscs.Trends Genet. 2020 Mar;36(3):189-202. doi: 10.1016/j.tig.2019.12.003. Epub 2020 Jan 14. Trends Genet. 2020. PMID: 31952839 Review.
Cited by
-
Patterns of gene flow define species of thermophilic Archaea.PLoS Biol. 2012 Feb;10(2):e1001265. doi: 10.1371/journal.pbio.1001265. Epub 2012 Feb 21. PLoS Biol. 2012. PMID: 22363207 Free PMC article.
-
Enantiomorphs and taxonomy of three conchological species in flat-shelled snails Trichocathaica (Pulmonata, Camaenidae).Zookeys. 2018 Dec 20;(810):19-44. doi: 10.3897/zookeys.810.29824. eCollection 2018. Zookeys. 2018. PMID: 30613172 Free PMC article.
-
Dual control by a single gene of secondary sexual characters and mating preferences in medaka.BMC Biol. 2009 Sep 29;7:64. doi: 10.1186/1741-7007-7-64. BMC Biol. 2009. PMID: 19788724 Free PMC article.
-
A speciation gene for left-right reversal in snails results in anti-predator adaptation.Nat Commun. 2010;1:133. doi: 10.1038/ncomms1133. Nat Commun. 2010. PMID: 21139578 Free PMC article.
-
Mouth asymmetry in the textbook example of scale-eating cichlid fish is not a discrete dimorphism after all.Proc Biol Sci. 2012 Dec 7;279(1748):4715-23. doi: 10.1098/rspb.2012.2082. Epub 2012 Oct 10. Proc Biol Sci. 2012. PMID: 23055070 Free PMC article.
References
-
- Brown NA, Wolpert L. The development of handedness in left/right asymmetry. Development. 1990;109:1–9. - PubMed
-
- McManus C. Right hand left hand. The origins of asymmetry in brains, bodies, atoms and cultures. London: Weidenfeld and Nicolson; 2002. 412 pp.
-
- Nonaka S, Shiratori H, Saijoh Y, Hamada H. Determination of left-right patterning of the mouse embryo by artificial nodal flow. Nature. 2002;418:96–99. - PubMed
-
- Bergmann DC, Lee M, Robertson B, Tsou MFB, Rose LS. Embryonic handedness choice in C. elegans involves the G alpha protein GPA-16. Development. 2003;130:5731–5740. - PubMed
-
- Shibazaki Y, Shimizu M, Kuroda R. Body handedness is directed by genetically determined cytoskeletal dynamics in the early embryo. Curr Biol. 2004;14:1462–1467. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
