

Role for a P-type H+-ATPase in the acidification of the endocytic pathway of Trypanosoma cruzi
- PMID: 16149915
- PMCID: PMC1316285
- DOI: 10.1042/BJ20051319
Role for a P-type H+-ATPase in the acidification of the endocytic pathway of Trypanosoma cruzi
Abstract
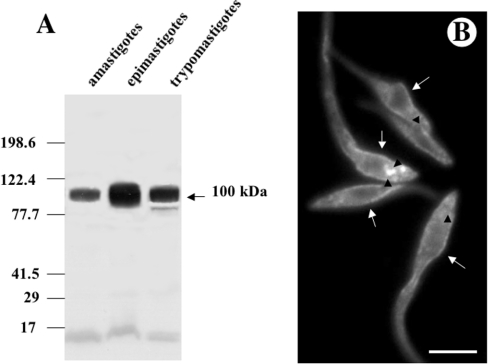
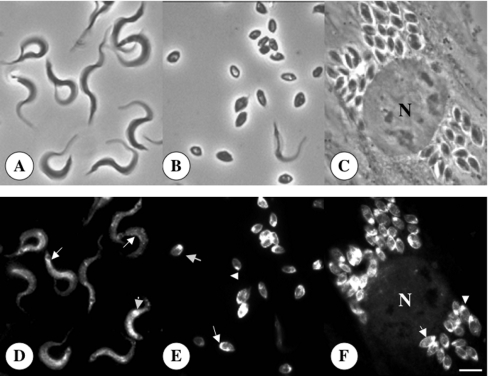
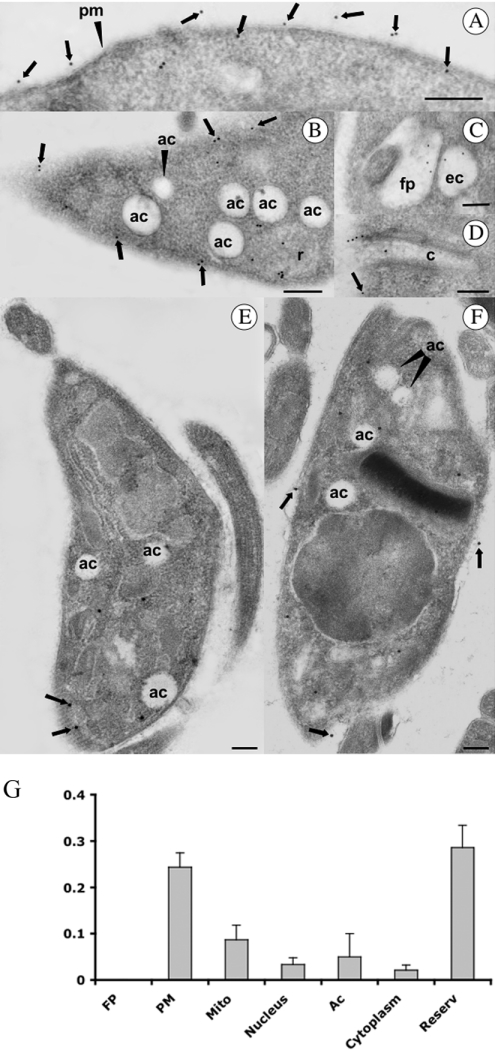
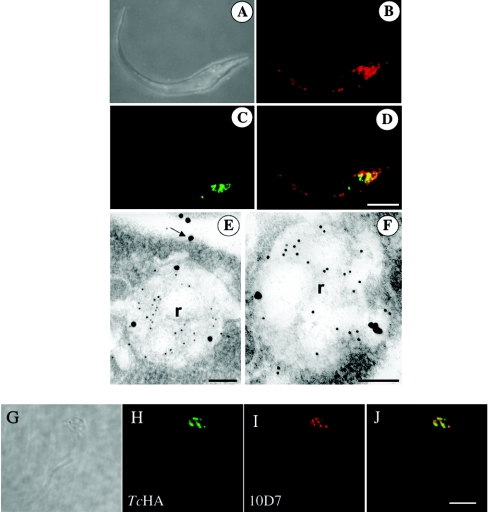
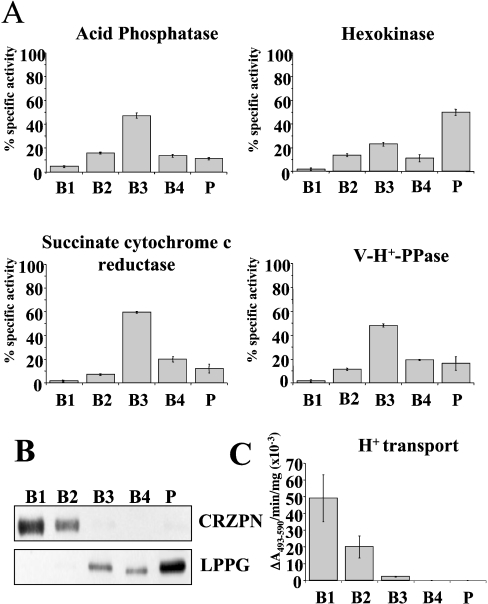
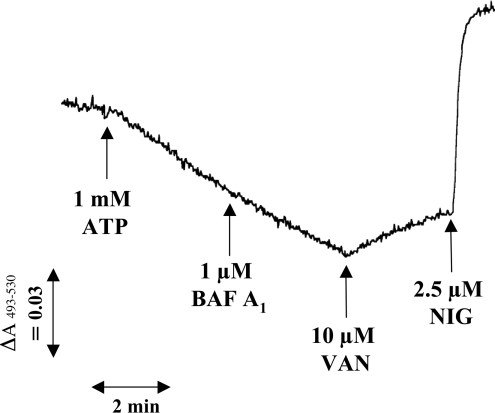
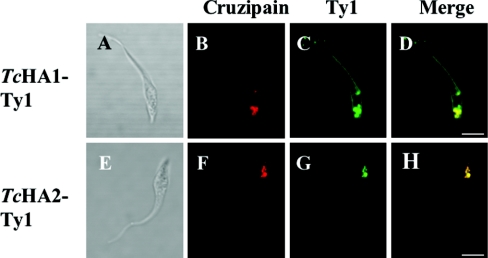
Previous studies in Trypanosoma cruzi, the etiologic agent of Chagas disease, have resulted in the cloning and sequencing of a pair of tandemly linked genes (TcHA1 and TcHA2) that encode P (phospho-intermediate form)-type H+-ATPases with homology to fungal and plant proton-pumping ATPases. In the present study, we demonstrate that these pumps are present in the plasma membrane and intracellular compartments of three different stages of T. cruzi. The main intracellular compartment containing these ATPases in epimastigotes was identified as the reservosome. This identification was achieved by immunofluorescence assays and immunoelectron microscopy showing their co-localization with cruzipain, and by subcellular fractionation and detection of their activity. ATP-dependent proton transport by isolated reservosomes was sensitive to vanadate and insensitive to bafilomycin A1, which is in agreement with the localization of P-type H+-ATPases in these organelles. Analysis by confocal immunofluorescence microscopy revealed that epitope-tagged TcHA1-Ty1 and TcHA2-Ty1 gene products are localized in the reservosomes, whereas the TcHA1-Ty1 gene product is additionally present in the plasma membrane. Immunogold electron microscopy showed the presence of the H+-ATPases in other compartments of the endocytic pathway such as the cytostome and endosomal vesicles, suggesting that in contrast with most cells investigated until now, the endocytic pathway of T. cruzi is acidified by a P-type H+-ATPase.
Figures
References
-
- Urbina J. A., Docampo R. Specific chemotherapy of Chagas disease: controversies and advances. Trends Parasitol. 2003;19:495–501. - PubMed
-
- Luo S., Scott D. A., Docampo R. Trypanosoma cruzi H+-ATPase 1 (TcHA1) and 2 (TcHA2) genes complement yeast mutants defective in H+ pumps and encode plasma membrane P-type H+-ATPases with different enzymatic properties. J. Biol. Chem. 2002;277:44497–44506. - PubMed
-
- Axelsen K. B., Palmgren M. G. Evolution of substrate specificities in the P-type ATPase superfamily. J. Mol. Evol. 1998;46:84–101. - PubMed
-
- Møller J. V., Juul B., le Maire M. Structural organization, ion transport, and energy transduction of P-type ATPases. Biochim. Biophys. Acta. 1996;1286:1–51. - PubMed
-
- Monk B. C., Perlin D. S. Fungal plasma membrane proton pumps as promising new antifungal targets. Crit. Rev. Microbiol. 1994;20:209–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources