

Dehydroepiandrosterone restoration of growth hormone gene expression in aging female rats, in vivo and in vitro: evidence for actions via estrogen receptors
- PMID: 16150906
- PMCID: PMC1868401
- DOI: 10.1210/en.2005-0811
Dehydroepiandrosterone restoration of growth hormone gene expression in aging female rats, in vivo and in vitro: evidence for actions via estrogen receptors
Abstract
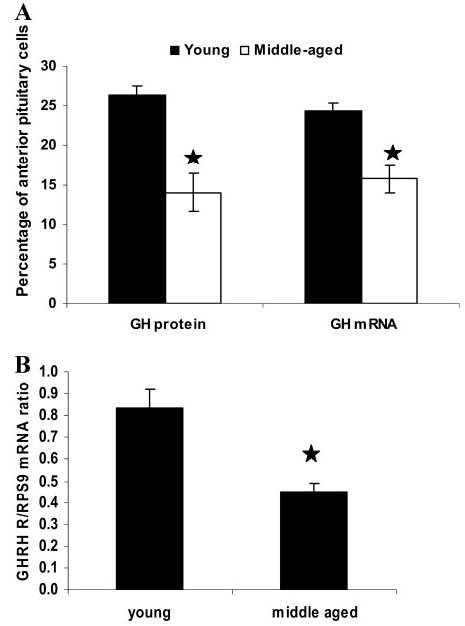
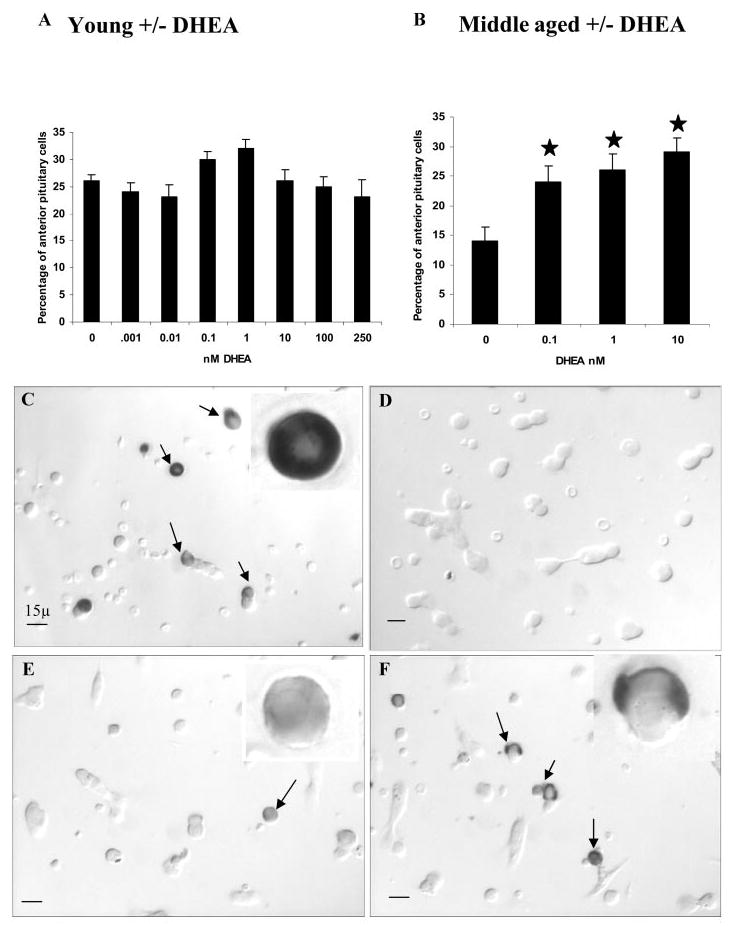
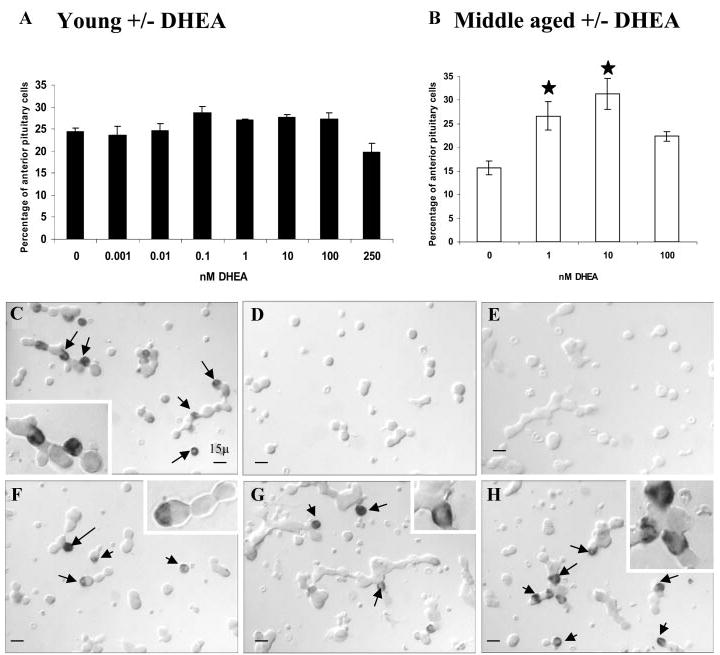
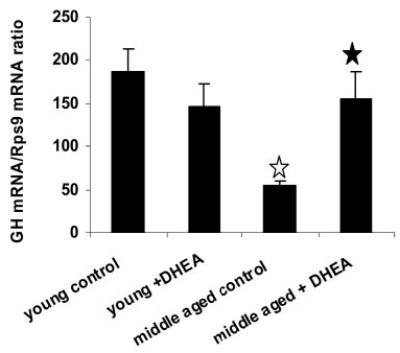
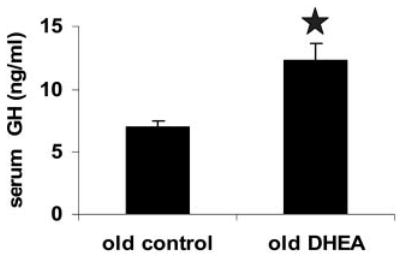
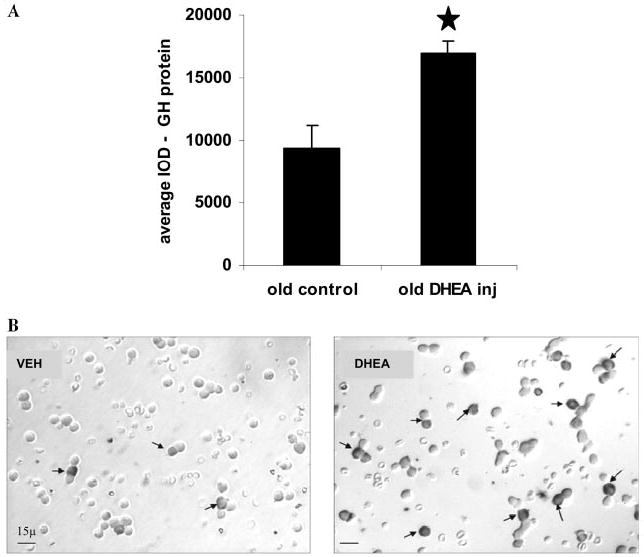
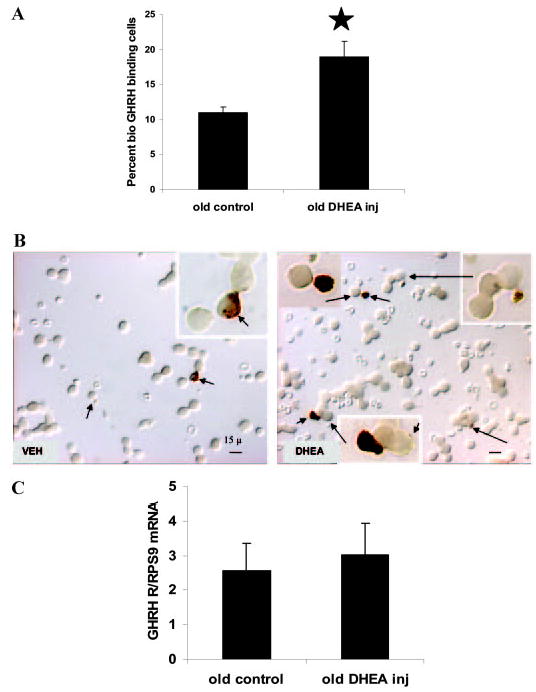
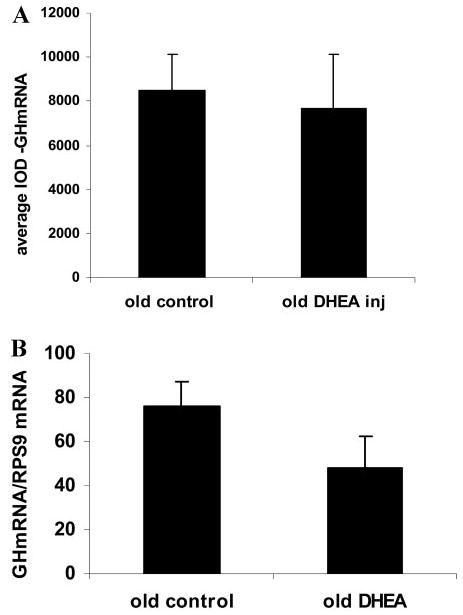
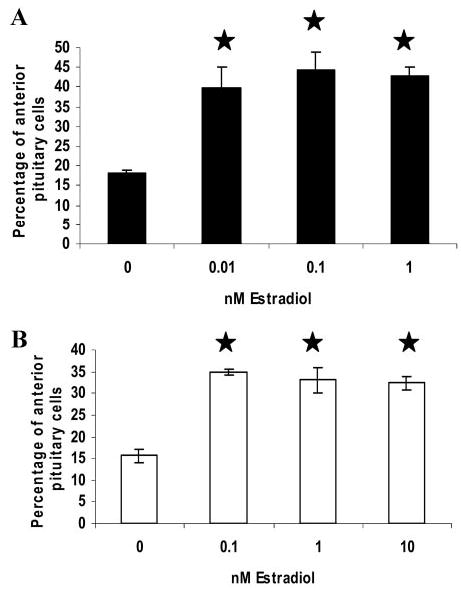
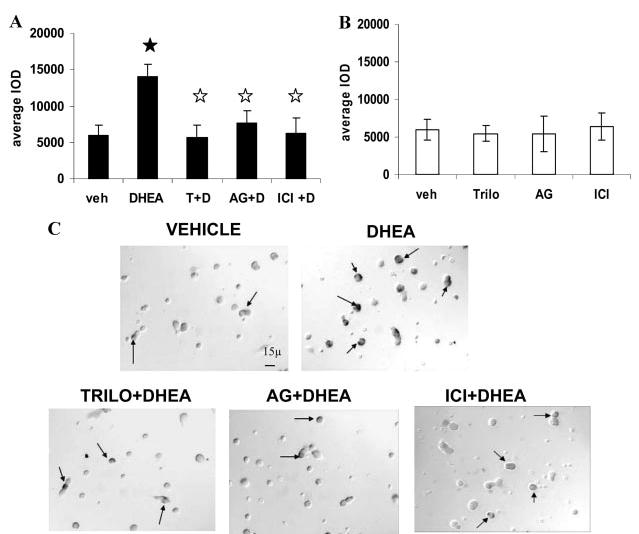
A decline in dehydroepiandrosterone (DHEA) and GH levels with aging may be associated with frailty and morbidity. Little is known about the direct effects of DHEA on somatotropes. We recently reported that 17beta-estradiol (E2), a DHEA metabolite, stimulates the expression of GH in vitro in young female rats. To test the hypothesis that DHEA restores function in aging somatotropes, dispersed anterior pituitary (AP) cells from middle-aged (12-14 months) or young (3-4 months) female rats were cultured in vitro with or without DHEA or E2 and fixed for immunolabeling or in situ hybridization. E2 increased the percentage of AP cells with GH protein or mRNA in the aged rats to young levels. DHEA increased the percentages of somatotropes (detected by GH protein or mRNA) from 14-16 +/- 2% to 29-31 +/- 3% (P < or = 0.05) and of GH mRNA (detected by quantitative RT-PCR) only in aging rats. To test DHEA's in vivo effects, 18-month-old female rats were injected with DHEA or vehicle for 2.5 d, followed by a bolus of GHRH 1 h before death. DHEA treatment increased serum GH 1.8-fold (7 +/- 0.5 to 12 +/- 1.3 ng/ml; P = 0.02, by RIA) along with a similar increase (P = 0.02) in GH immunolabel. GHRH target cells also increased from 11 +/- 1% to 19 +/- 2% (P = 0.03). Neither GH nor GHRH receptor mRNAs levels were changed. To test the mechanisms behind DHEA's actions, AP cells from aging rats were treated with DHEA with or without inhibitors of DHEA metabolism. Trilostane, aminogluthemide, or ICI 182,780 completely blocked the stimulatory effects of DHEA, suggesting that DHEA metabolites may stimulate aging somatotropes via estrogen receptors.
Figures
Similar articles
-
Functional modification of pituitary somatotropes in the aromatase knockout mouse and the effect of estrogen replacement.Endocrinology. 2004 Feb;145(2):604-12. doi: 10.1210/en.2003-0646. Epub 2003 Oct 16. Endocrinology. 2004. PMID: 14563698
-
Oestrogen replacement in vivo rescues the dysfunction of pituitary somatotropes in ovariectomised aromatase knockout mice.Neuroendocrinology. 2005;81(3):158-66. doi: 10.1159/000086864. Epub 2005 Jul 5. Neuroendocrinology. 2005. PMID: 16015026
-
Effect of GHRH and GHRP-2 treatment in vitro on GH secretion and levels of GH, pituitary transcription factor-1, GHRH-receptor, GH-secretagogue-receptor and somatostatin receptor mRNAs in ovine pituitary cells.Eur J Endocrinol. 2004 Feb;150(2):235-42. doi: 10.1530/eje.0.1500235. Eur J Endocrinol. 2004. PMID: 14763922
-
Structure and regulation of expression of the mouse GH receptor.J Endocrinol. 2002 Oct;175(1):55-9. doi: 10.1677/joe.0.1750055. J Endocrinol. 2002. PMID: 12379490 Review.
-
The actions of exogenous dehydroepiandrosterone in experimental animals and humans.Proc Soc Exp Biol Med. 1998 Jul;218(3):174-91. doi: 10.3181/00379727-218-44285. Proc Soc Exp Biol Med. 1998. PMID: 9648935 Review.
Cited by
-
The somatotrope as a metabolic sensor: deletion of leptin receptors causes obesity.Endocrinology. 2011 Jan;152(1):69-81. doi: 10.1210/en.2010-0498. Epub 2010 Nov 17. Endocrinology. 2011. PMID: 21084451 Free PMC article.
-
Tissue-Specific Effects of Loss of Estrogen during Menopause and Aging.Front Endocrinol (Lausanne). 2012 Feb 8;3:19. doi: 10.3389/fendo.2012.00019. eCollection 2012. Front Endocrinol (Lausanne). 2012. PMID: 22654856 Free PMC article.
-
The Effects of Neuroactive Steroids on Myelin in Health and Disease.Med Princ Pract. 2024;33(3):198-214. doi: 10.1159/000537794. Epub 2024 Feb 13. Med Princ Pract. 2024. PMID: 38350432 Free PMC article. Review.
-
Age-Related Changes in MicroRNA in the Rat Pituitary and Potential Role in GH Regulation.Int J Mol Sci. 2018 Jul 15;19(7):2058. doi: 10.3390/ijms19072058. Int J Mol Sci. 2018. PMID: 30011963 Free PMC article.
-
Ablation of leptin signaling to somatotropes: changes in metabolic factors that cause obesity.Endocrinology. 2012 Oct;153(10):4705-15. doi: 10.1210/en.2012-1331. Epub 2012 Aug 3. Endocrinology. 2012. PMID: 22865370 Free PMC article.
References
-
- Wilshire GB, Loughlin JS, Brown JR, Adel TE, Santoro N. Diminished function of the somatotropic axis in older reproductive-aged women. J Clin Endocrinol Metab. 1995;80:608–613. - PubMed
-
- Toogood AA. Growth hormone (GH) status and body composition in normal ageing and in elderly adults with GH deficiency. Horm Res. 2003;60:105–111. - PubMed
-
- Lamberts SW, van den Beld AW, van der Lely AJ. The endocrinology of aging. Science. 1997;278:419–424. - PubMed
-
- Ezzat S, Fear S, Gaillard RC, Gayle C, Landy H, Marcovitz S, Mattioni T, Nussey S, Rees A, Svanberg E. Gender-specific responses of lean body composition and non-gender-specific cardiac function improvement after GH replacement in GH-deficient adults. J Clin Endocrinol Metab. 2002;87:2725–2733. - PubMed
-
- Attanasio AF, Bates PC, Ho KK, Webb SM, Ross RJ, Strasburger CJ, Bouillon R, Crowe B, Selander K, Valle D, Lamberts SW. Human growth hormone replacement in adult hypopituitary patients: long-term effects on body composition and lipid status–3-year results from the HypoCCS database. J Clin Endocrinol Metab. 2002;87:1600–1606. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
