

The radioresistance kinase TLK1B protects the cells by promoting repair of double strand breaks
- PMID: 16156902
- PMCID: PMC1242231
- DOI: 10.1186/1471-2199-6-19
The radioresistance kinase TLK1B protects the cells by promoting repair of double strand breaks
Abstract
Background: The mammalian protein kinase TLK1 is a homologue of Tousled, a gene involved in flower development in Arabidopsis thaliana. The function of TLK1 is not well known, although knockout of the gene in Drosophila or expression of a dominant negative mutant in mouse cells causes loss of nuclear divisions and missegregation of chromosomes probably, due to alterations in chromatin remodeling capacity. Overexpression of TLK1B, a spliced variant of the TLK1 mRNA, in a model mouse cell line increases it's resistance to ionizing radiation (IR) or the radiomimetic drug doxorubicin, also likely due to changes in chromatin remodeling. TLK1B is translationally regulated by the availability of the translation factor eIF4E, and its synthesis is activated by IR. The reason for this mechanism of regulation is likely to provide a rapid means of promoting repair of DSBs. TLK1B specifically phosphorylates histone H3 and Asf1, likely resulting in changes in chromatin structure, particularly at double strand breaks (DSB) sites.
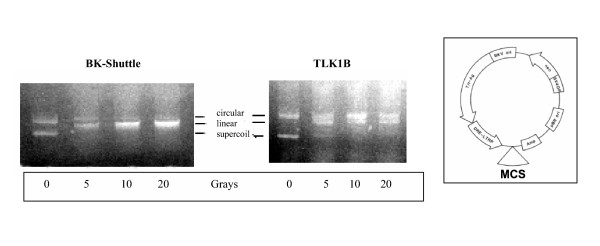
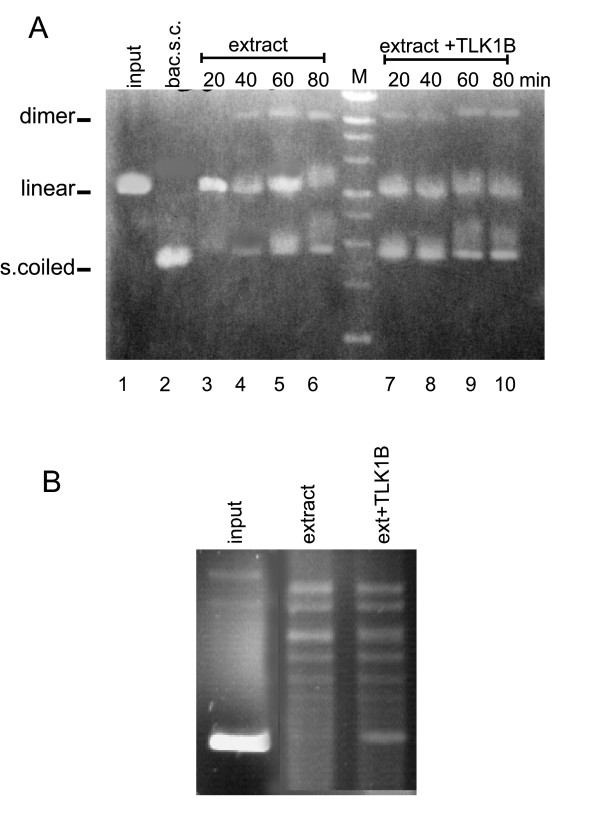
Results: In this work, we provide several lines of evidence that TLK1B protects the cells from IR by facilitating the repair of DSBs. First, the pattern of phosphorylation and dephosphorylation of H2AX and H3 indicated that cells overexpressing TLK1B return to pre-IR steady state much more rapidly than controls. Second, the repair of episomes damaged with DSBs was much more rapid in cells overexpressing TLK1B. This was also true for repair of genomic damage. Lastly, we demonstrate with an in vitro repair system that the addition of recombinant TLK1B promotes repair of a linearized plasmid incubated with nuclear extract. In addition, TLK1B in this in vitro system promotes the assembly of chromatin as shown by the formation of more highly supercoiled topomers of the plasmid.
Conclusion: In this work, we provide evidence that TLK1B promotes the repair of DSBs, likely as a consequence of a change in chromatin remodeling capacity that must precede the assembly of repair complexes at the sites of damage.
Figures





Similar articles
-
TLK1B promotes repair of UV-damaged DNA through chromatin remodeling by Asf1.BMC Mol Biol. 2006 Oct 20;7:37. doi: 10.1186/1471-2199-7-37. BMC Mol Biol. 2006. PMID: 17054786 Free PMC article.
-
Translation of the radioresistance kinase TLK1B is induced by gamma-irradiation through activation of mTOR and phosphorylation of 4E-BP1.BMC Mol Biol. 2004 Apr 7;5:1. doi: 10.1186/1471-2199-5-1. BMC Mol Biol. 2004. PMID: 15070431 Free PMC article.
-
Tousled homolog, TLK1, binds and phosphorylates Rad9; TLK1 acts as a molecular chaperone in DNA repair.DNA Repair (Amst). 2009 Jan 1;8(1):87-102. doi: 10.1016/j.dnarep.2008.09.005. Epub 2008 Nov 5. DNA Repair (Amst). 2009. PMID: 18940270
-
Preserving salivary gland physiology against genotoxic damage - the Tousled way.Oral Dis. 2018 Nov;24(8):1390-1398. doi: 10.1111/odi.12836. Epub 2018 May 2. Oral Dis. 2018. PMID: 29383801 Free PMC article. Review.
-
Chromatin modification and NBS1: their relationship in DNA double-strand break repair.Genes Genet Syst. 2016;90(4):195-208. doi: 10.1266/ggs.15-00010. Epub 2015 Nov 25. Genes Genet Syst. 2016. PMID: 26616756 Review.
Cited by
-
TLK1B mediated phosphorylation of Rad9 regulates its nuclear/cytoplasmic localization and cell cycle checkpoint.BMC Mol Biol. 2016 Feb 9;17:3. doi: 10.1186/s12867-016-0056-x. BMC Mol Biol. 2016. PMID: 26860083 Free PMC article.
-
Differential requirements for Tousled-like kinases 1 and 2 in mammalian development.Cell Death Differ. 2017 Nov;24(11):1872-1885. doi: 10.1038/cdd.2017.108. Epub 2017 Jul 14. Cell Death Differ. 2017. PMID: 28708136 Free PMC article.
-
Phenothiazine Inhibitors of TLKs Affect Double-Strand Break Repair and DNA Damage Response Recovery and Potentiate Tumor Killing with Radiomimetic Therapy.Genes Cancer. 2013 Jan;4(1-2):39-53. doi: 10.1177/1947601913479020. Genes Cancer. 2013. PMID: 23946870 Free PMC article.
-
High yield bacterial expression, purification and characterisation of bioactive Human Tousled-like Kinase 1B involved in cancer.Sci Rep. 2018 Mar 19;8(1):4796. doi: 10.1038/s41598-018-22744-5. Sci Rep. 2018. PMID: 29555908 Free PMC article.
-
TLK1 as a therapeutic target in TMZ resistant glioblastoma using small molecule inhibitor.Sci Rep. 2025 Apr 26;15(1):14691. doi: 10.1038/s41598-025-86599-3. Sci Rep. 2025. PMID: 40287404 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
