

A novel mechanism of actin filament processive capping by formin: solution of the rotation paradox
- PMID: 16157699
- PMCID: PMC2171452
- DOI: 10.1083/jcb.200504156
A novel mechanism of actin filament processive capping by formin: solution of the rotation paradox
Abstract
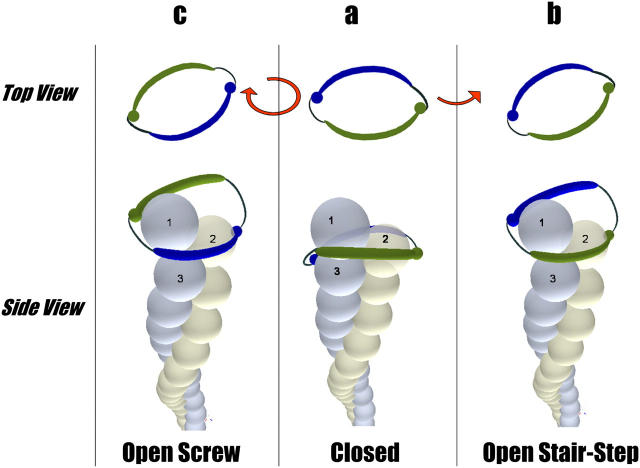
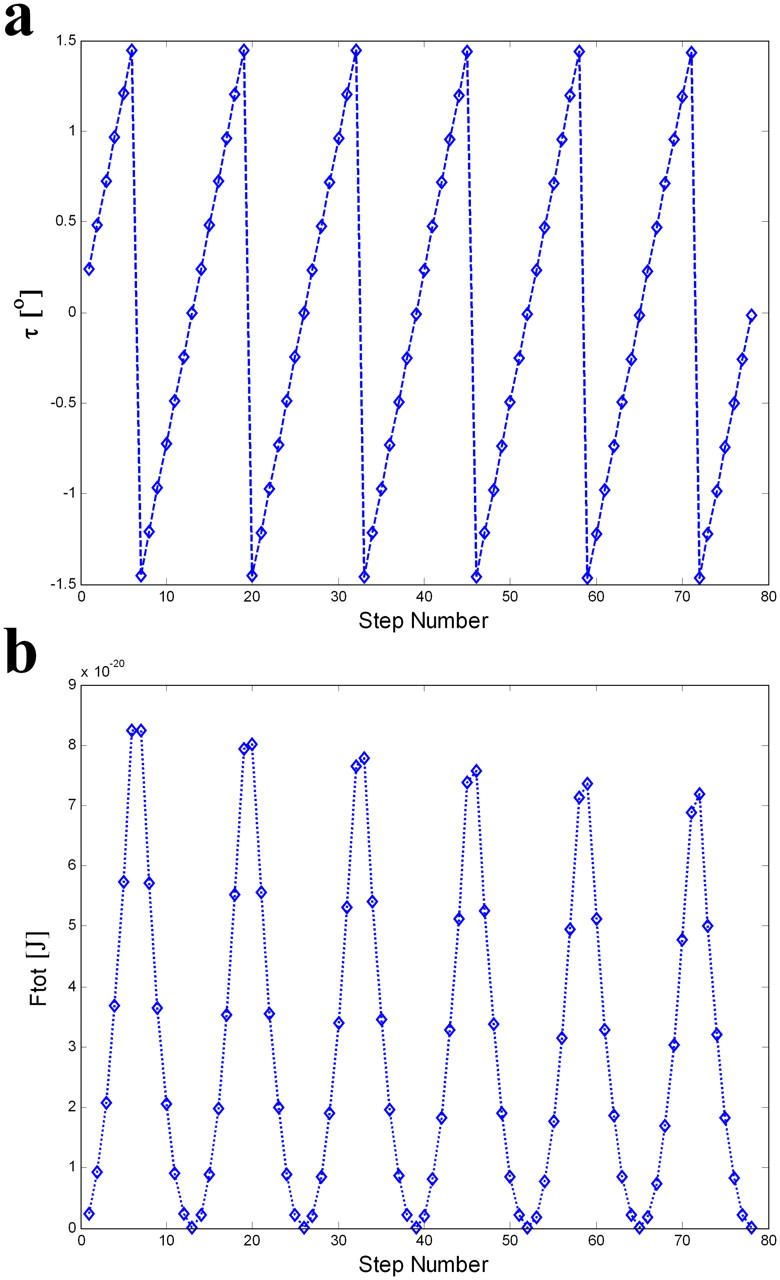
The FH2 domains of formin family proteins act as processive cappers of actin filaments. Previously suggested stair-stepping mechanisms of processive capping imply that a formin cap rotates persistently in one direction with respect to the filament. This challenges the formin-mediated mechanism of intracellular cable formation. We suggest a novel scenario of processive capping that is driven by developing and relaxing torsion elastic stresses. Based on the recently discovered crystal structure of an FH2-actin complex, we propose a second mode of processive capping-the screw mode. Within the screw mode, the formin dimer rotates with respect to the actin filament in the direction opposite to that generated by the stair-stepping mode so that a combination of the two modes prevents persistent torsion strain accumulation. We determine an optimal regime of processive capping, whose essence is a periodic switch between the stair-stepping and screw modes. In this regime, elastic energy does not exceed feasible values, and supercoiling of actin filaments is prevented.
Figures
References
-
- Higashida, C., T. Miyoshi, A. Fujita, F. Oceguera-Yanez, J. Monypenny, Y. Andou, S. Narumiya, and N. Watanabe. 2004. Actin polymerization-driven molecular movement of mDia1 in living cells. Science. 303:2007–2010. - PubMed
-
- Higgs, H.N. 2005. Formin proteins: a domain-based approach. Trends Biochem. Sci. 30:342–353. - PubMed
-
- Holmes, K.C., D. Popp, W. Gebhard, and W. Kabsch. 1990. Atomic model of the actin filament. Nature. 347:44–49. - PubMed
-
- Isambert, H., P. Venier, A.C. Maggs, A. Fattoum, R. Kassab, D. Pantaloni, and M.F. Carlier. 1995. Flexibility of actin filaments derived from thermal fluctuations. Effect of bound nucleotide, phalloidin, and muscle regulatory proteins. J. Biol. Chem. 270:11437–11444. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
