

Snail induction is an early response to Gli1 that determines the efficiency of epithelial transformation
- PMID: 16158046
- PMCID: PMC1361531
- DOI: 10.1038/sj.onc.1209077
Snail induction is an early response to Gli1 that determines the efficiency of epithelial transformation
Abstract
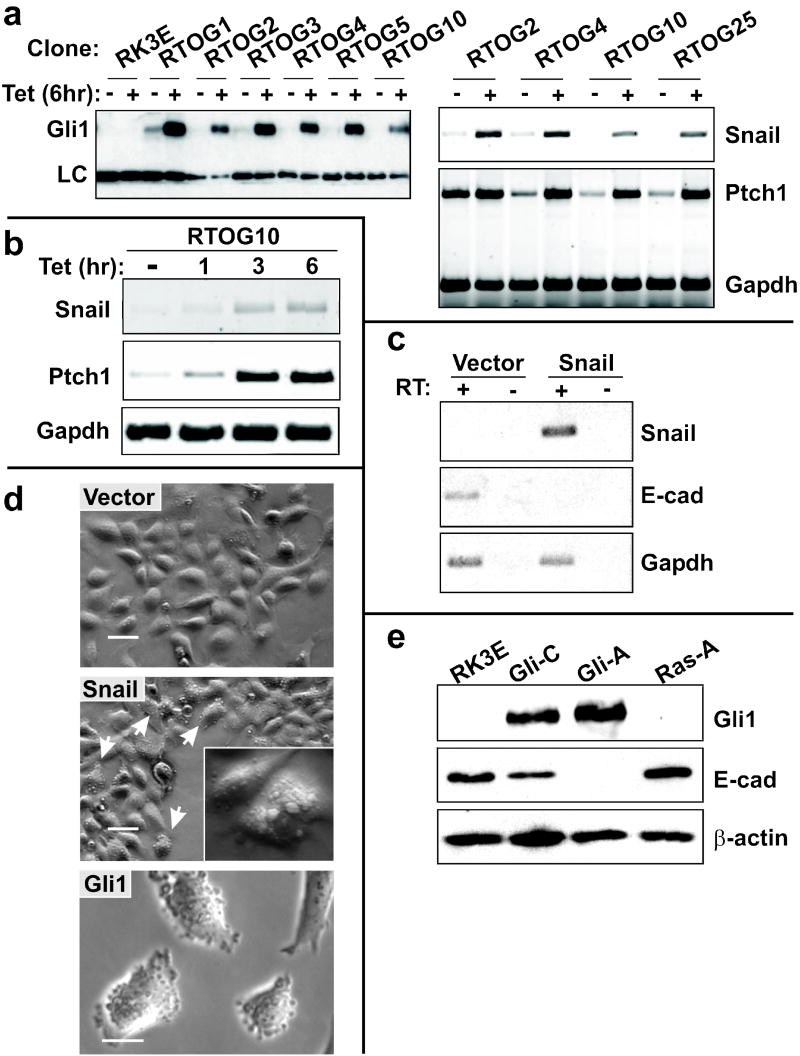
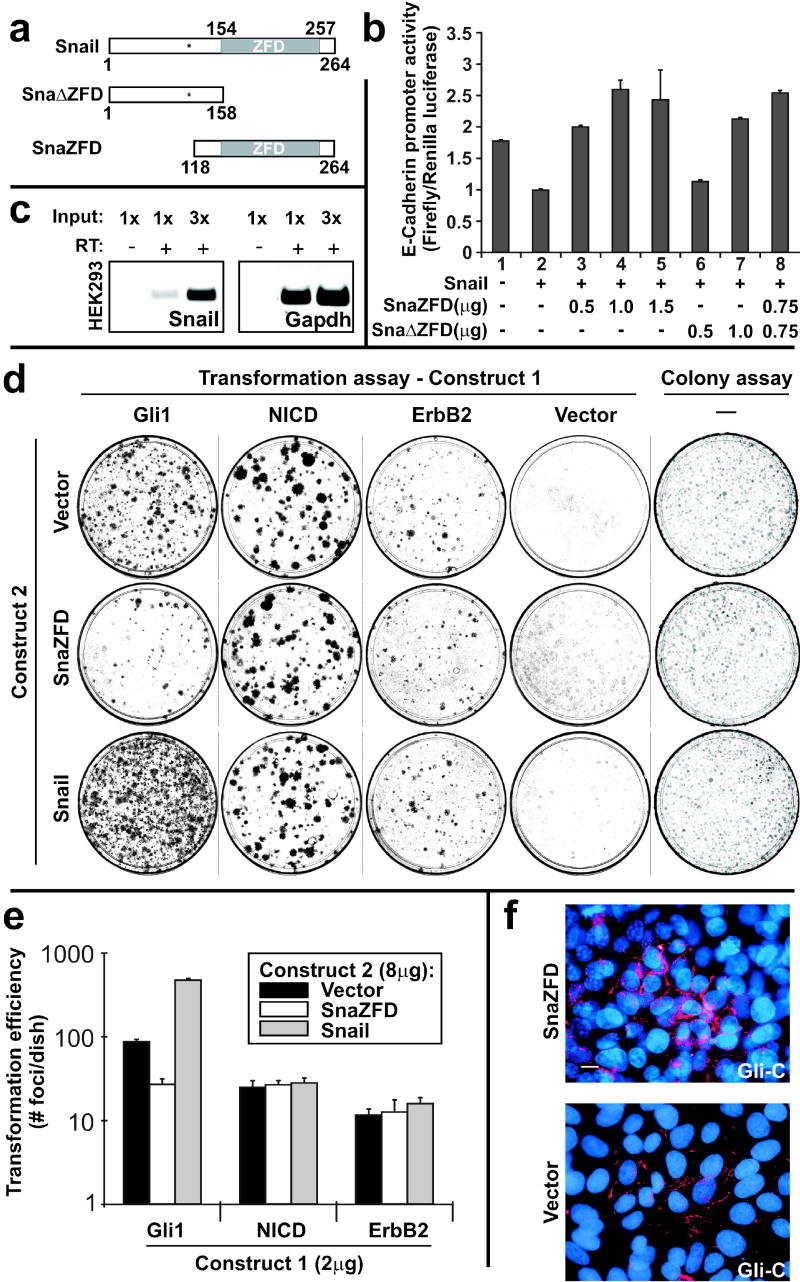
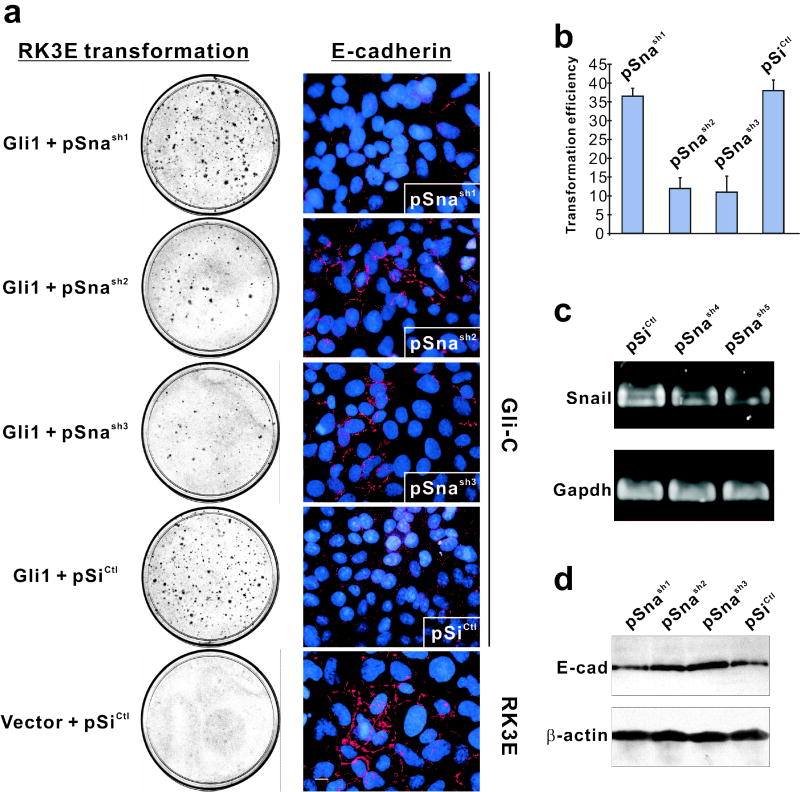
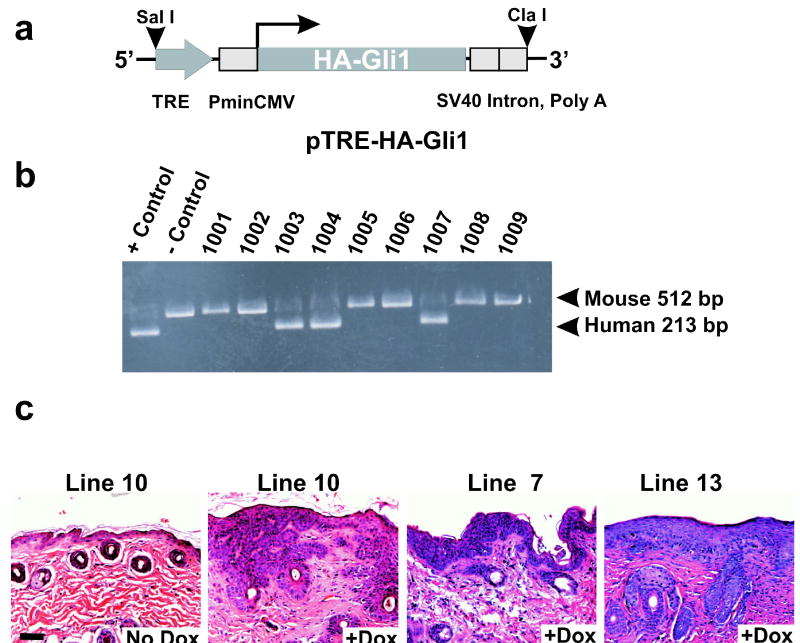
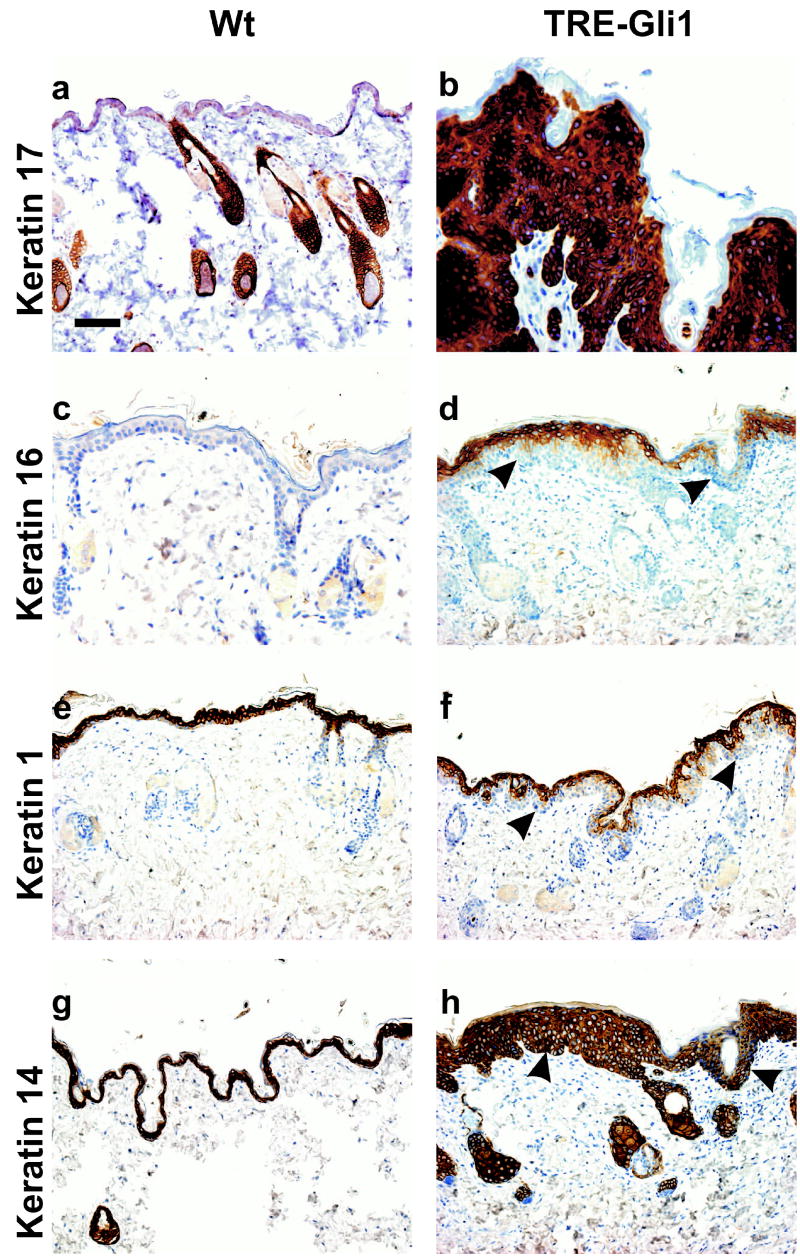
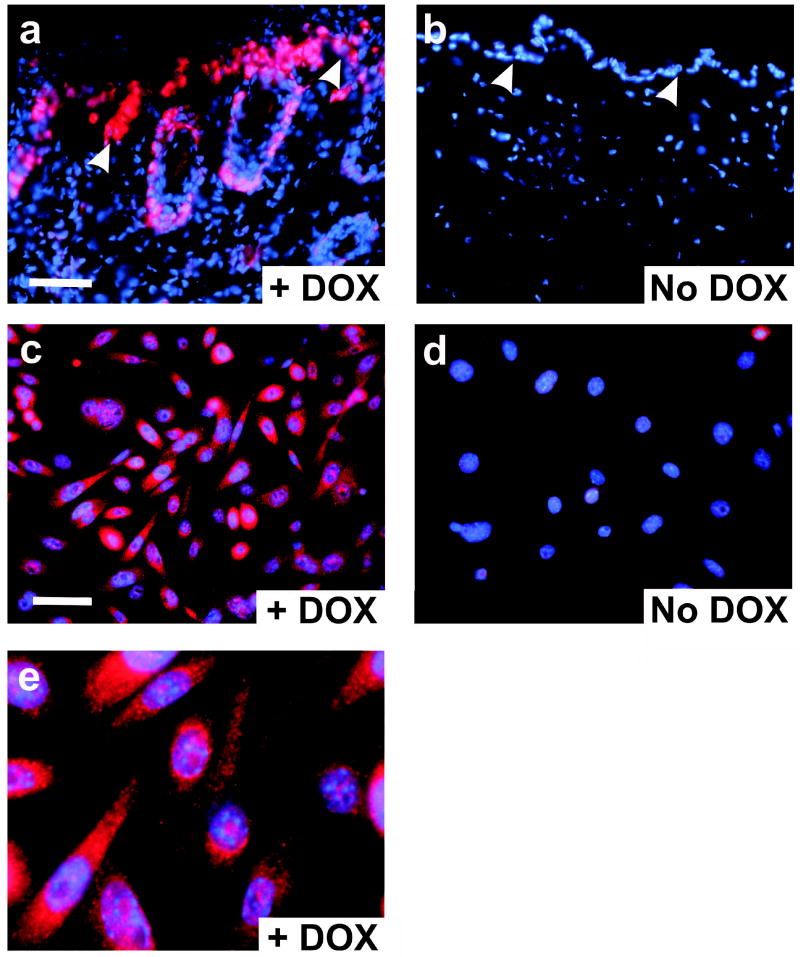
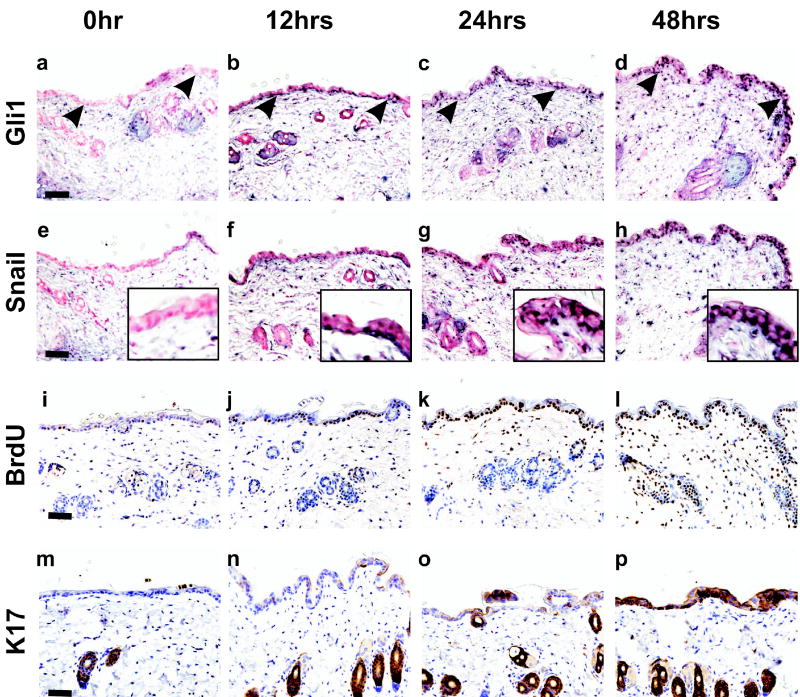
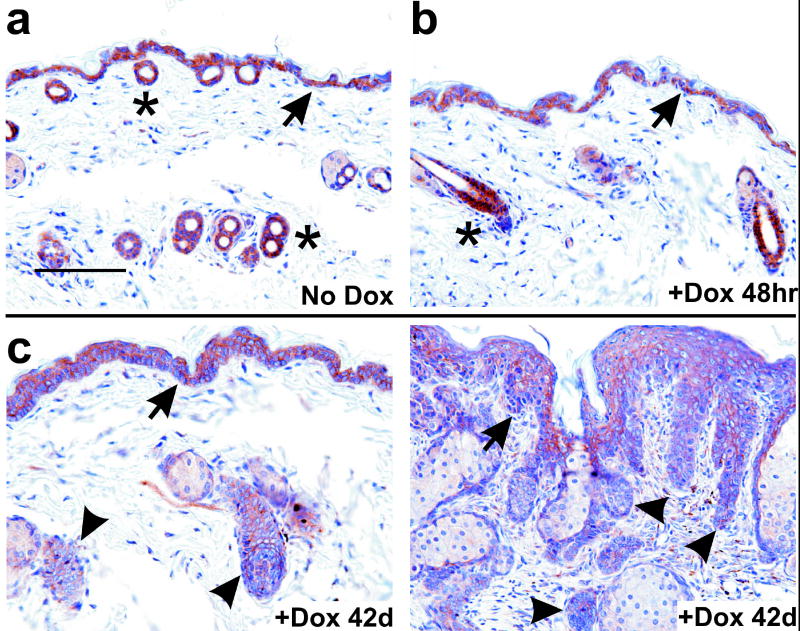
Gli family members mediate constitutive Hedgehog signaling in the common skin cancer, basal cell carcinoma (BCC). Snail/Snai1 is rapidly induced by Gli1 in vitro, and is coexpressed with Gli1 in human hair follicles and skin tumors. In the current study, we generated a dominant-negative allele of Snail, SnaZFD, composed of the zinc-finger domain and flanking sequence. In promoter-reporter assays, SnaZFD blocked the activity of wild-type Snail on the E-cadherin promoter. Snail loss-of-function mediated by SnaZFD or by one of several short hairpin RNAs inhibited transformation of RK3E epithelial cells by Gli1. Conversely, enforced expression of Snail promoted transformation in vitro by Gli1, but not by other genes that were tested, including Notch1, ErbB2, and N-Ras. As observed for Gli1, wild-type Snail repressed E-cadherin in RK3E cells and induced blebbing of the cytoplasmic membrane. Induction of a conditional Gli1 transgene in the basal keratinocytes of mouse skin led to rapid upregulation of Snail transcripts and to cell proliferation in the interfollicular epidermis. Established Gli1-induced skin lesions exhibited molecular similarities to BCC, including loss of E-cadherin. The results identify Snail as a Gli1-inducible effector of transformation in vitro, and an early Gli1-responsive gene in the skin.
Figures
References
-
- Agren M, Kogerman P, Kleman MI, Wessling M, Toftgard R. Gene. 2004;330:101–14. 101–114. - PubMed
-
- Ascano JM, Beverly LJ, Capobianco AJ. J Biol Chem. 2003;278:8771–8779. - PubMed
-
- Aszterbaum M, Epstein J, Oro A, Douglas V, LeBoit PE, Scott MP, Epstein EH., Jr Nat Med. 1999;5:1285–1291. - PubMed
-
- Aybar MJ, Nieto MA, Mayor R. Development. 2003;130:483–494. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
