

Contributions of domesticated plant studies to our understanding of plant evolution
- PMID: 16159942
- PMCID: PMC4247096
- DOI: 10.1093/aob/mci259
Contributions of domesticated plant studies to our understanding of plant evolution
Abstract
Background: Plant evolutionary theory has been greatly enriched by studies on crop species. Over the last century, important information has been generated on many aspects of population biology, speciation and polyploid genetics.
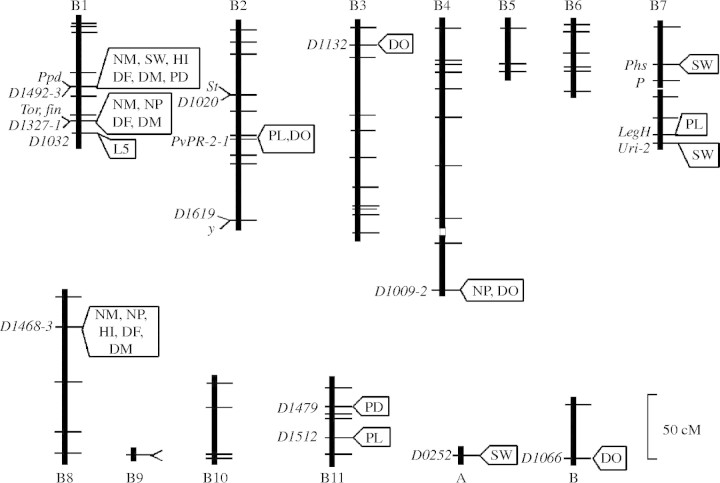
Scope: Searches for quantitative trait loci (QTL) in crop species have uncovered numerous blocks of genes that have dramatic effects on adaptation, particularly during the domestication process. Many of these QTL have epistatic and pleiotropic effects making rapid evolutionary change possible. Most of the pioneering work on the molecular basis of self-incompatibility has been conducted on crop species, along with the sequencing of the phytopathogenic resistance genes (R genes) responsible for the 'gene-to-gene' relations of coevolution observed in host-pathogen relationships. Some of the better examples of co-adaptation and early acting inbreeding depression have also been elucidated in crops. Crop-wild progenitor interactions have provided rich opportunities to study the evolution of novel adaptations subsequent to hybridization. Most crop/wild F1 hybrids have reduced fitness, but in some instances the crop relatives have acquired genes that make them more efficient weeds through crop mimicry. Studies on autopolyploid alfalfa and potato have uncovered the means by which polyploid gametes are formed and have led to hypotheses about how multiallelic interactions are associated with fitness and self-fertility. Research on the cole crops and wheat has discovered that newly formed polyploids can undergo dramatic genome rearrangements that could lead to rapid evolutionary change.
Conclusions: Many more important evolutionary discoveries are on the horizon, now that the whole genome sequence is available of the two major subspecies of rice Oryza sativa ssp. japonica and O. sativa ssp. indica. The rice sequence data can be used to study the origin of genes and gene families, track rates of sequence divergence over time, and provide hints about how genes evolve and generate products with novel biological properties. The rice sequence data has already been mined to show that transposable elements often carry fragments of cellular genes. This type of genome shuffling could play a role in creating novel, reorganized genes with new adaptive properties.
Figures
References
-
- Allard RW. 1988. Genetic changes associated with the evolution of adaptedness in cultivated plants and their wild progenitors. Journal of Heredity 79: 225–238. - PubMed
-
- Allard RW. 1990. The genetics of host–pathogen coevolution: implications for genetic resource conservation. Journal of Heredity 81: 1–6. - PubMed
-
- Armstead LP, Turner LB, Farrell M, Skot L, Gomez P, Montoya T, Donnison IS, King IP, Humphreys MO. 2004. Synteny between a major heading-date QTL in perennial ryegrass (Lolium perenne L.) and the Hd3 Heading-date locus in rice. Theoretical and Applied Genetics 108: 822–828. - PubMed
-
- Barber HN. 1970. Hybridization and the evolution of plants. Taxon 19: 154–160.
