

Endocytic processing of connexin43 gap junctions: a morphological study
- PMID: 16162097
- PMCID: PMC1383664
- DOI: 10.1042/BJ20050674
Endocytic processing of connexin43 gap junctions: a morphological study
Abstract
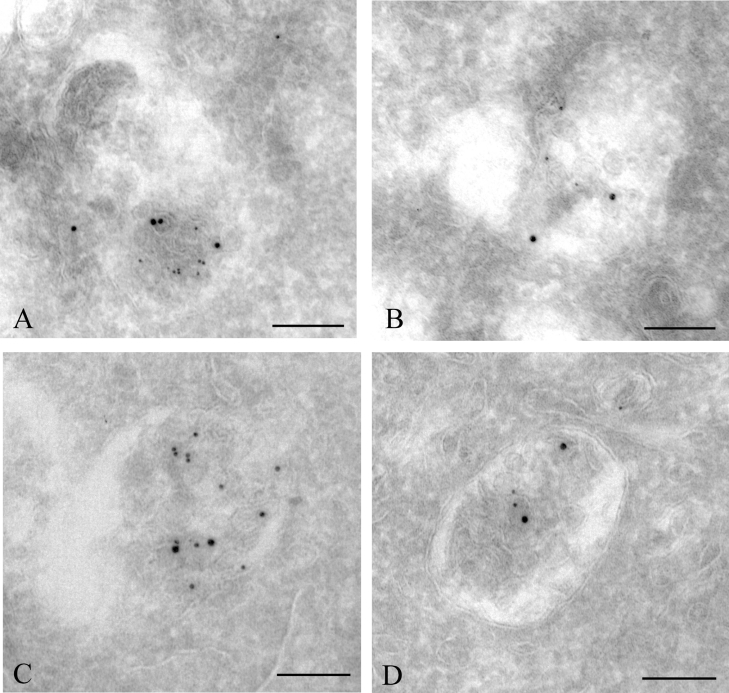
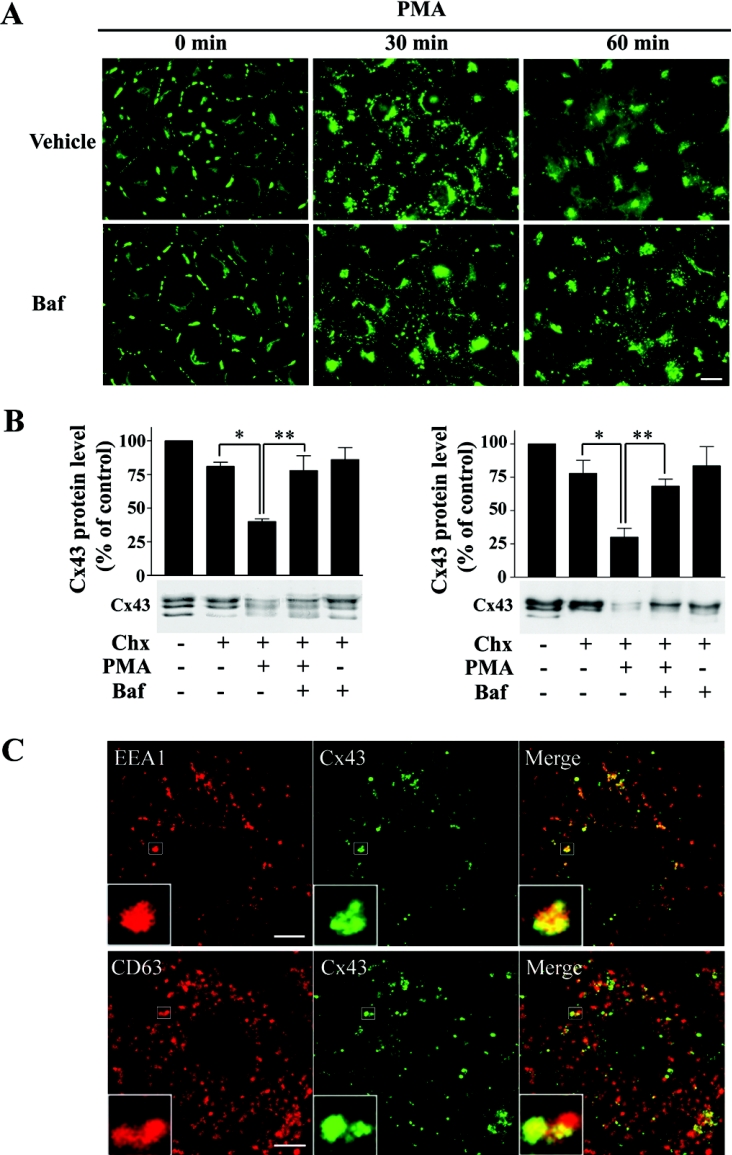
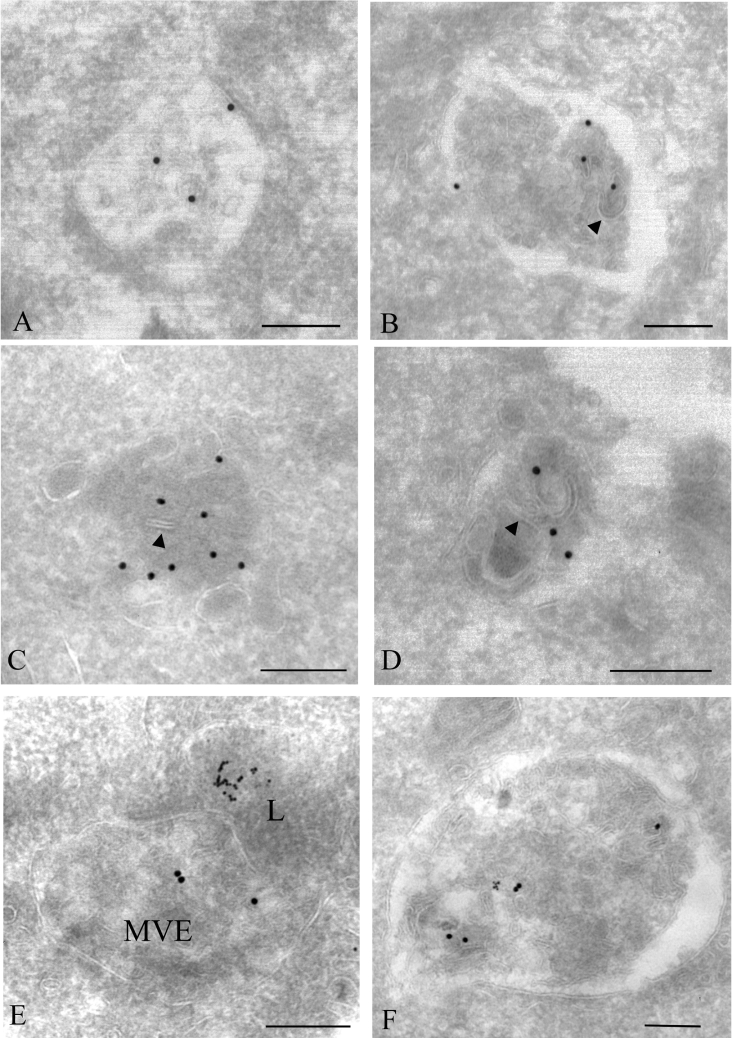
Gap junctions are plasma membrane areas enriched in channels that provide direct intercellular communication. Gap junctions have a high turnover rate; however, the mechanisms by which gap junctions are degraded are incompletely understood. In the present study, we show that in response to phorbol ester treatment, the gap junction channel protein Cx43 (connexin43) is redistributed from the plasma membrane to intracellular vesicles positive for markers for early and late endosomes and for the endolysosomal protease cathepsin D. Immunoelectron microscopy studies indicate that the double membranes of internalized gap junctions undergo separation and cutting, resulting in multivesicular endosomes enriched in Cx43 protein. Using preloading of BSA-gold conjugates to mark lysosomes, we provide evidence suggesting that the degradation process of the double-membrane structure of annular gap junctions occurs prior to transport of Cx43 to the lysosome. The results further suggest that bafilomycin A1, an inhibitor of vacuolar H+-ATPases, causes accumulation of Cx43 in early endosomes. Taken together, these findings indicate that internalized gap junctions undergo a maturation process from tightly sealed double-membrane vacuoles to connexin-enriched multivesicular endosomes with a single limiting membrane. The results further suggest that along with the processing of the double-membrane structure of annular gap junctions, connexins are trafficked via early and late endosomes, finally resulting in their endolysosomal degradation.
Figures
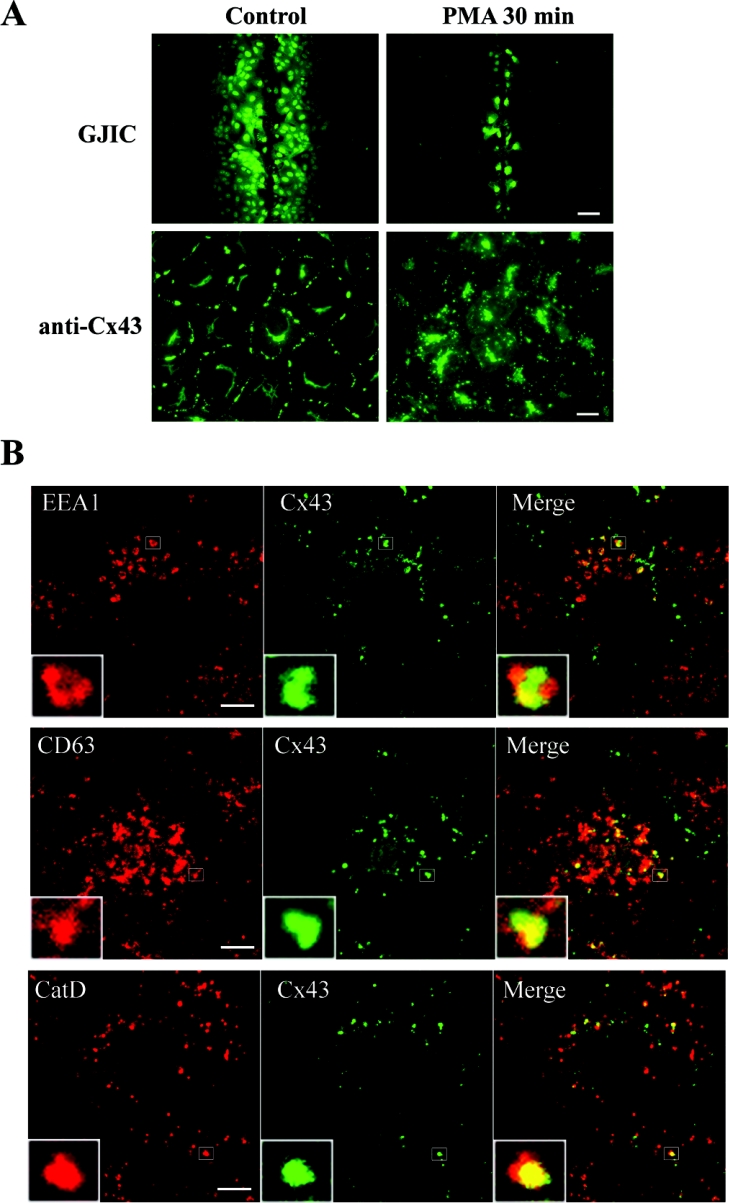
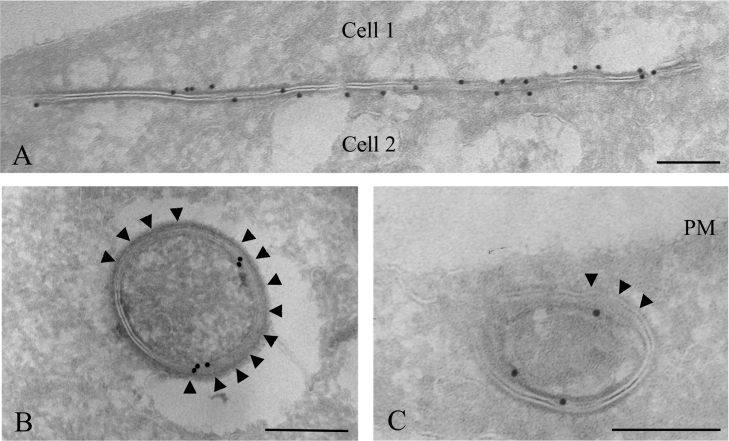
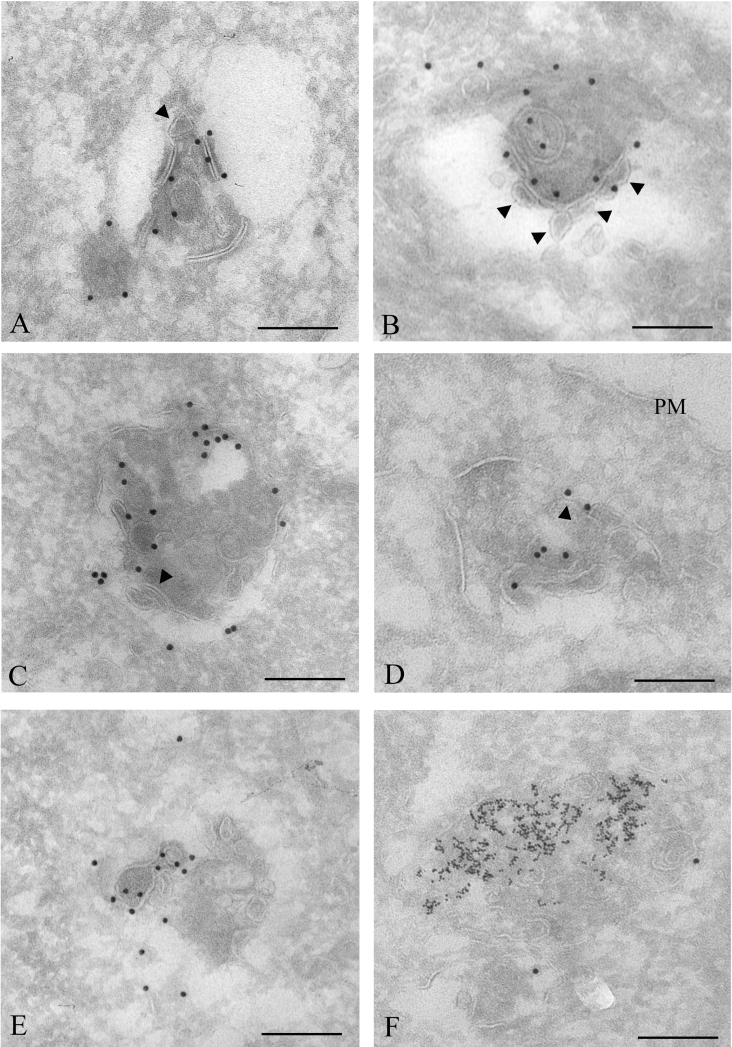
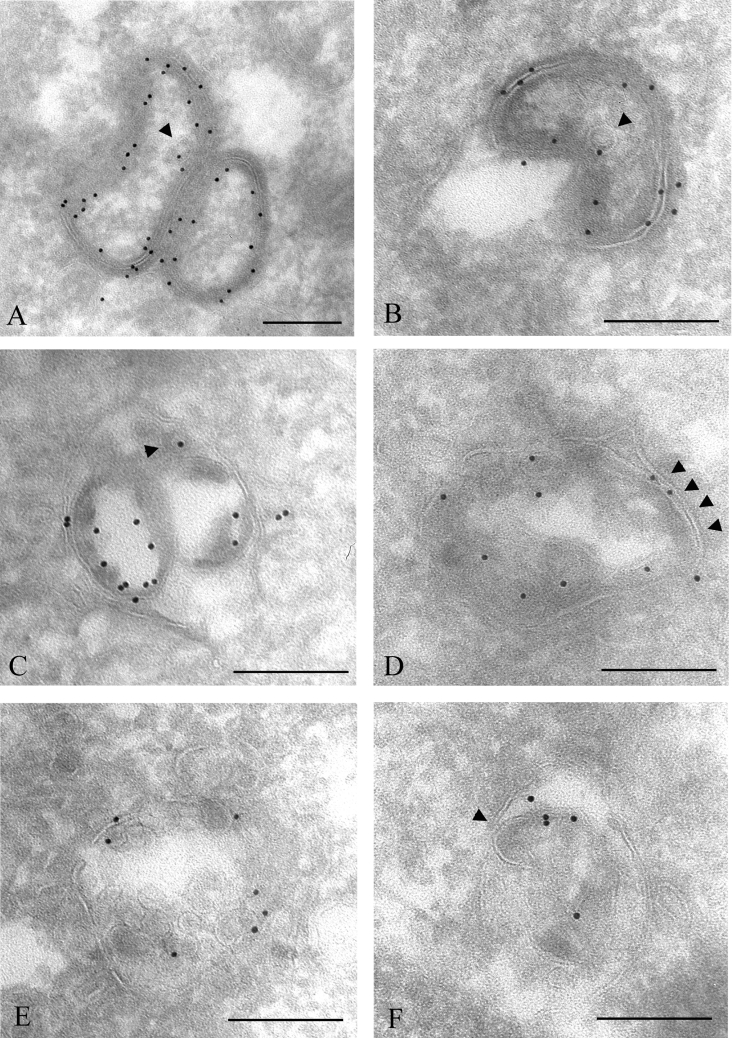
References
-
- Saez J. C., Berthoud V. M., Branes M. C., Martinez A. D., Beyer E. C. Plasma membrane channels formed by connexins: their regulation and functions. Physiol. Rev. 2003;83:1359–1400. - PubMed
-
- Musil L. S., Goodenough D. A. Multisubunit assembly of an integral plasma membrane channel protein, gap junction connexin43, occurs after exit from the ER. Cell (Cambridge, Mass.) 1993;74:1065–1077. - PubMed
-
- Sosinsky G. E. Molecular organization of gap junction membrane channels. J. Bioenerg. Biomembr. 1996;28:297–309. - PubMed
-
- Segretain D., Falk M. M. Regulation of connexin biosynthesis, assembly, gap junction formation, and removal. Biochim. Biophys. Acta. 2004;1662:3–21. - PubMed
-
- Wei C. J., Xu X., Lo C. W. Connexins and cell signaling in development and disease. Annu. Rev. Cell Dev. Biol. 2004;20:811–838. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous