

Dual-mode recognition of noncanonical tRNAs(Ser) by seryl-tRNA synthetase in mammalian mitochondria
- PMID: 16163389
- PMCID: PMC1276171
- DOI: 10.1038/sj.emboj.7600811
Dual-mode recognition of noncanonical tRNAs(Ser) by seryl-tRNA synthetase in mammalian mitochondria
Abstract
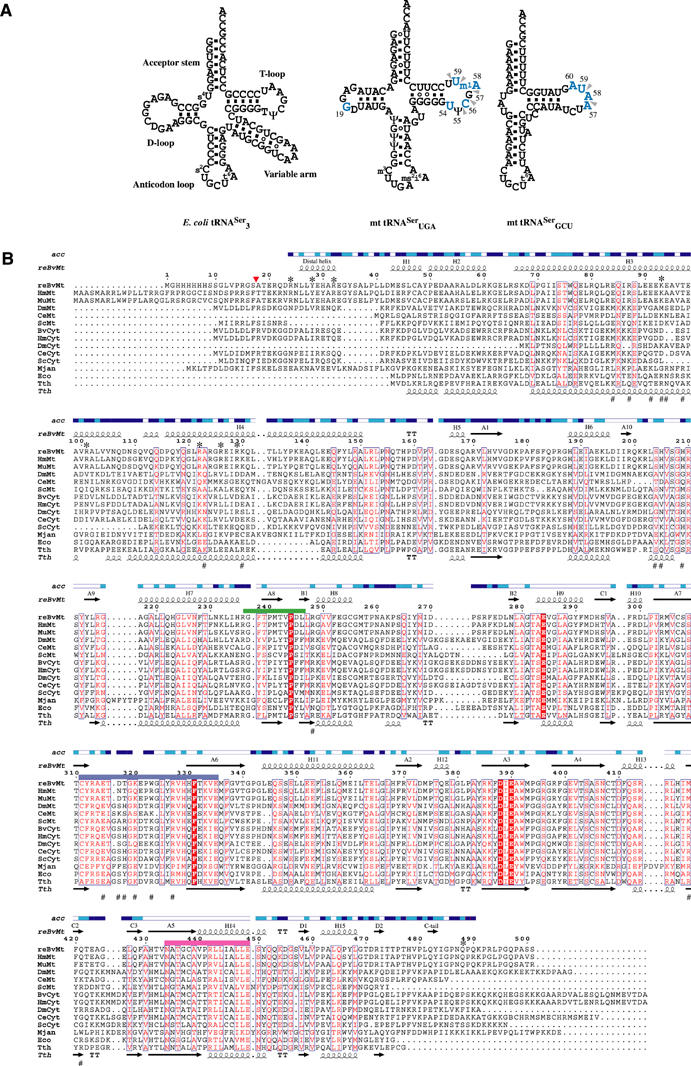
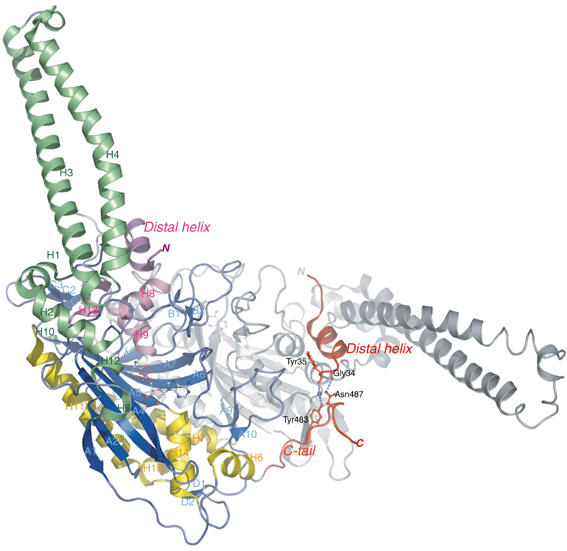
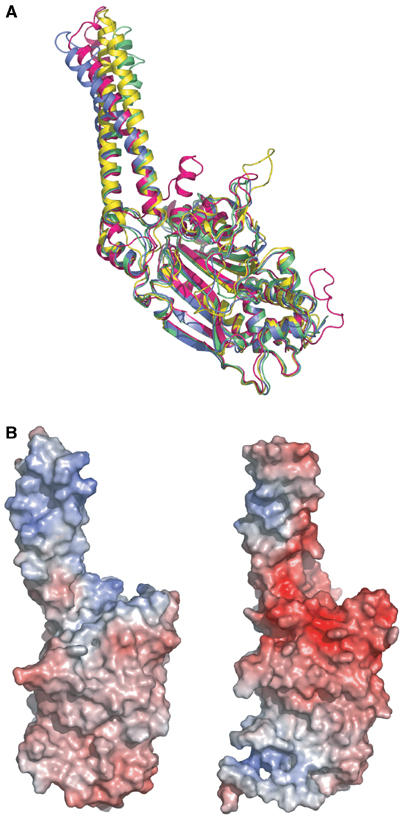
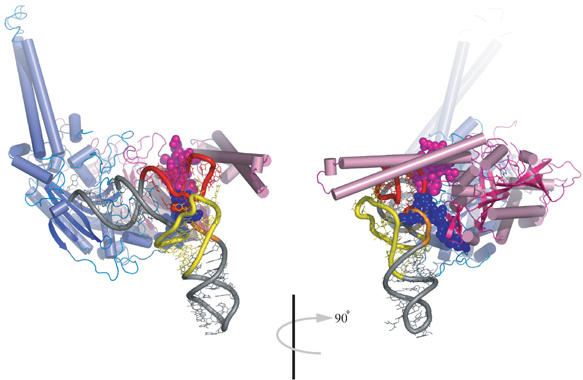
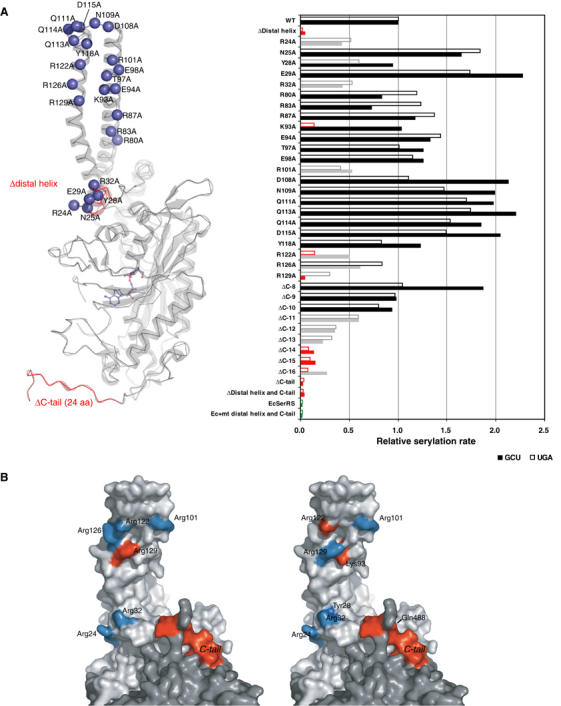
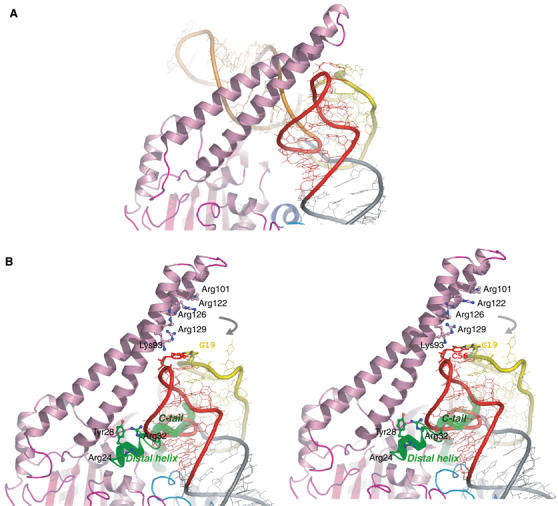
The secondary structures of metazoan mitochondrial (mt) tRNAs(Ser) deviate markedly from the paradigm of the canonical cloverleaf structure; particularly, tRNA(Ser)(GCU) corresponding to the AGY codon (Y=U and C) is highly truncated and intrinsically missing the entire dihydrouridine arm. None of the mt serine isoacceptors possesses the elongated variable arm, which is the universal landmark for recognition by seryl-tRNA synthetase (SerRS). Here, we report the crystal structure of mammalian mt SerRS from Bos taurus in complex with seryl adenylate at an atomic resolution of 1.65 A. Coupling structural information with a tRNA-docking model and the mutagenesis studies, we have unraveled the key elements that establish tRNA binding specificity, differ from all other known bacterial and eukaryotic systems, are the characteristic extensions in both extremities, as well as a few basic residues residing in the amino-terminal helical arm of mt SerRS. Our data further uncover an unprecedented mechanism of a dual-mode recognition employed to discriminate two distinct 'bizarre' mt tRNAs(Ser) by alternative combination of interaction sites.
Figures
References
-
- Belrhali H, Yaremchuk A, Tukalo M, Larsen K, Berthet-Colominas C, Leberman R, Beijer B, Sproat B, Als-Nielsen J, Grubel G, Legrand JF, Lehmann M, Cusack S (1994) Crystal structures at 2.5 angstrom resolution of seryl-tRNA synthetase complexed with two analogs of seryl adenylate. Science 263: 1432–1436 - PubMed
-
- Bilokapic S, Korencic D, Soll D, Weygand-Durasevic I (2004) The unusual methanogenic seryl-tRNA synthetase recognizes tRNASer species from all three kingdoms of life. Eur J Biochem 271: 694–702 - PubMed
-
- Biou V, Yaremchuk A, Tukalo M, Cusack S (1994) The 2.9 A crystal structure of T. thermophilus seryl-tRNA synthetase complexed with tRNASer. Science 263: 1404–1410 - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D 54 (Part 5): 905–921 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
