

Long-term in vivo modulation of synaptic efficacy at the neuromuscular junction of Rana pipiens frogs
- PMID: 16166159
- PMCID: PMC1464201
- DOI: 10.1113/jphysiol.2005.094805
Long-term in vivo modulation of synaptic efficacy at the neuromuscular junction of Rana pipiens frogs
Abstract
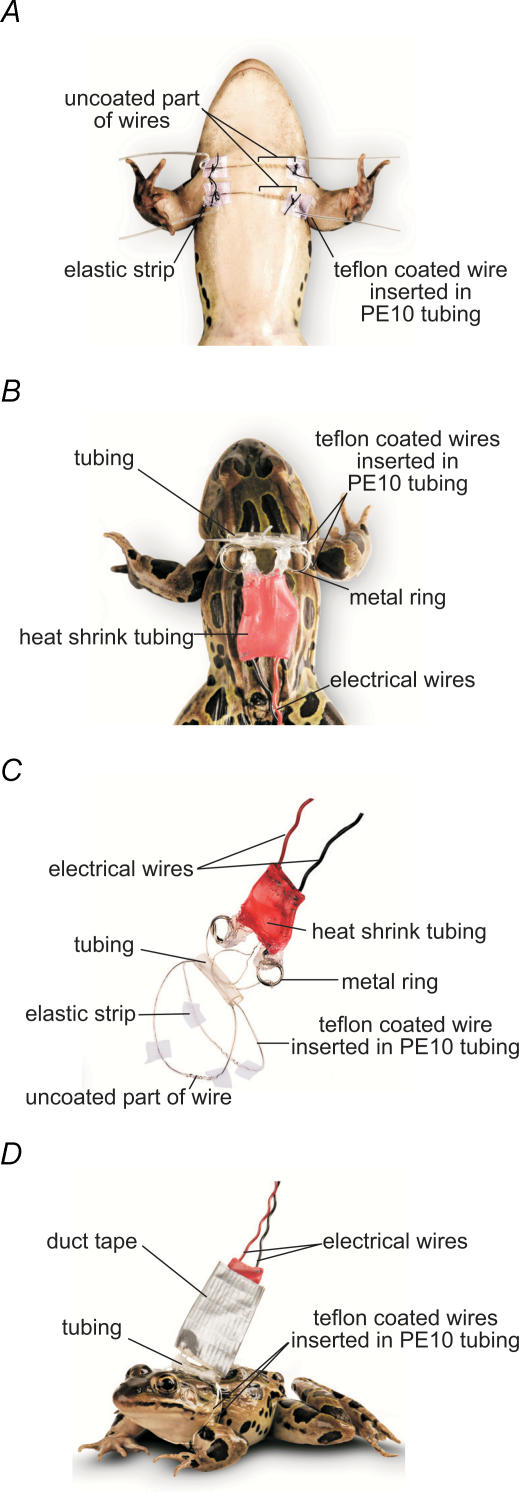
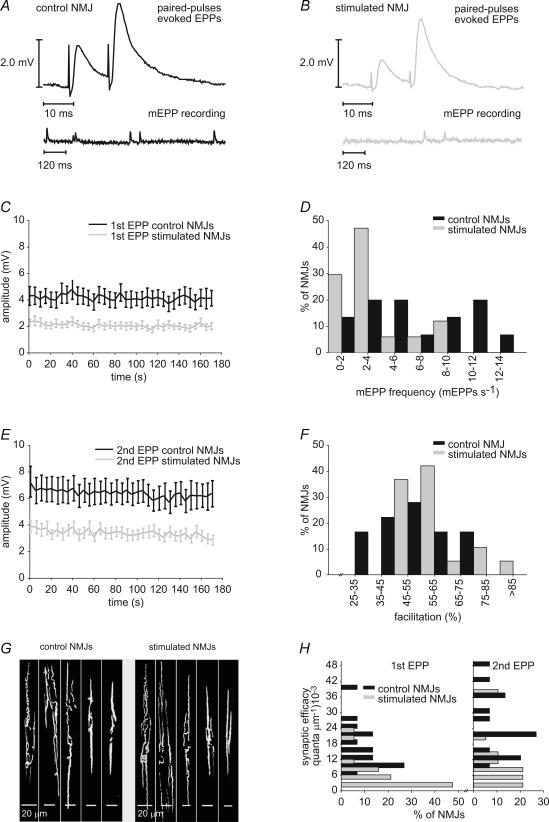
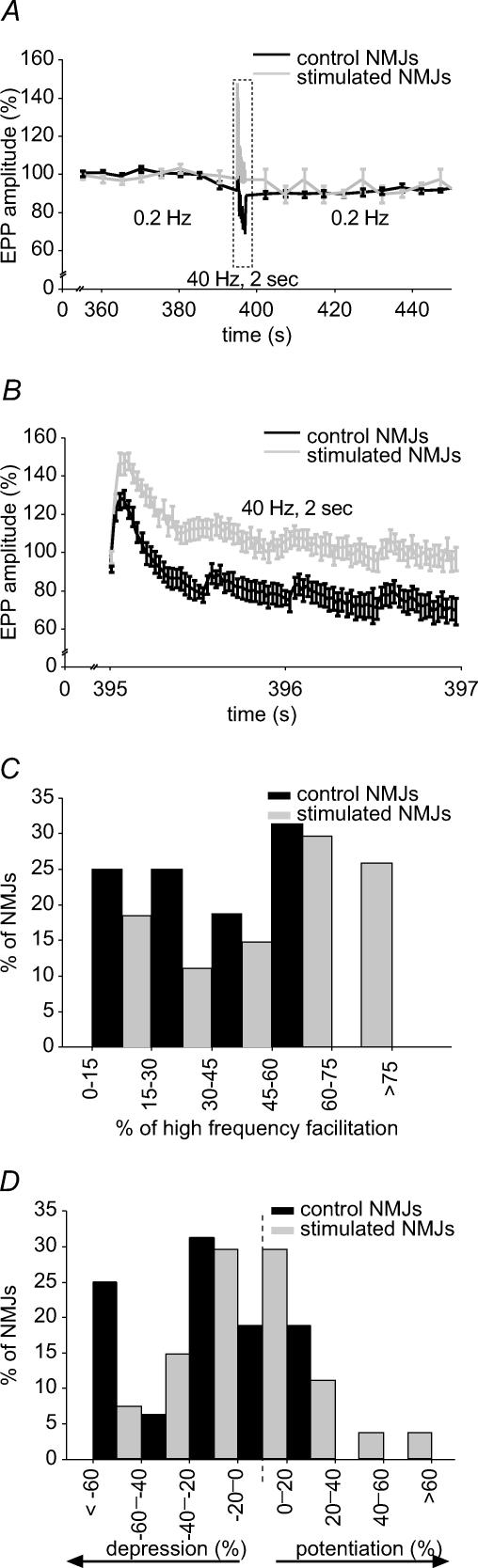
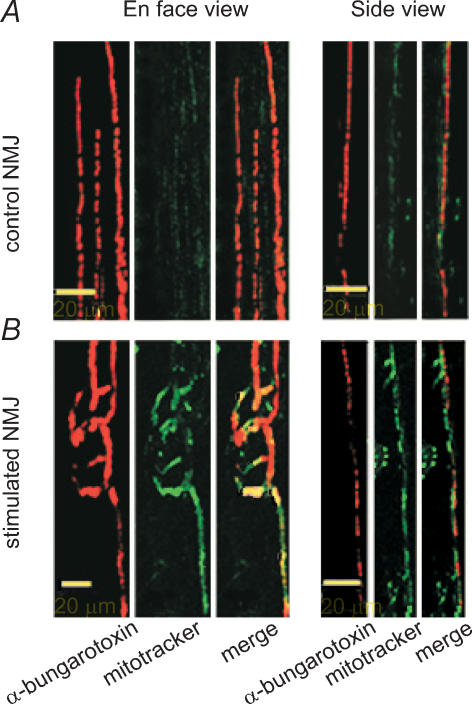
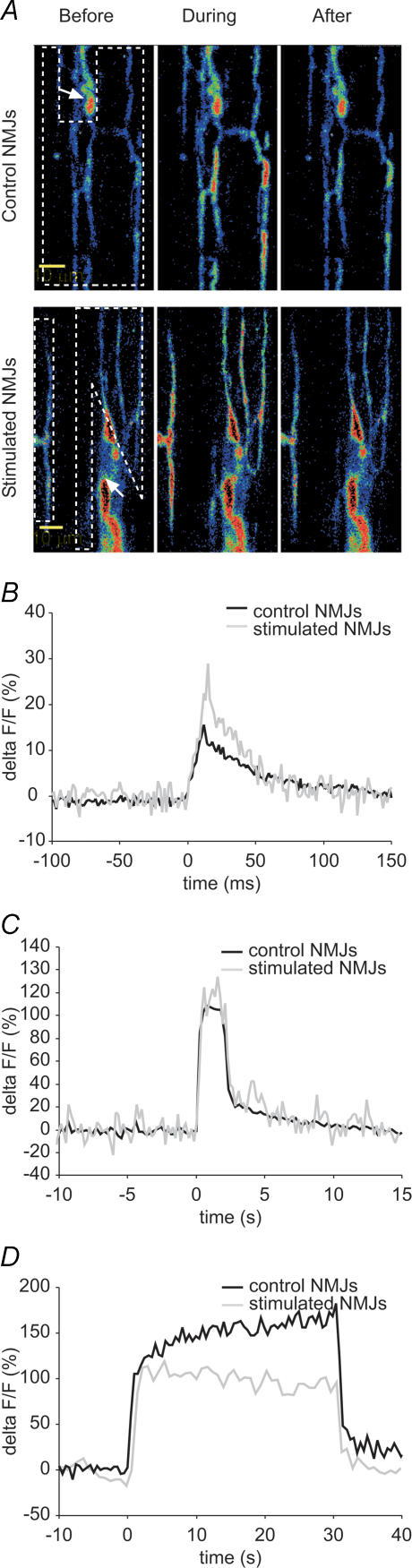
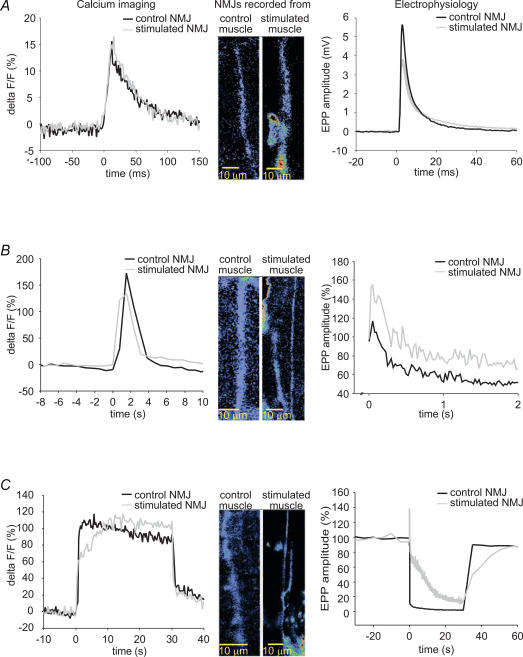
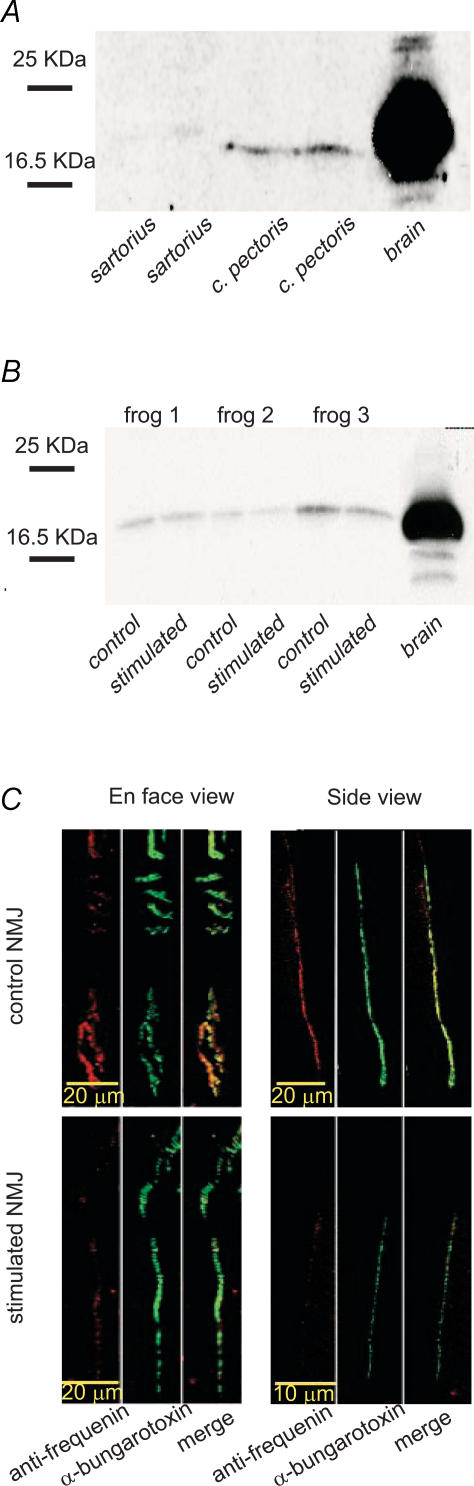
Prolonged changes in motor neurone activity can result in long-term changes in synaptic transmission. We investigated whether mechanisms commonly thought to be involved in determining synaptic efficacy of vertebrate motor neurones are involved in these long-term changes. The nerve supplying the cutaneous pectoris muscle was chronically stimulated via skin surface electrodes in freely moving frogs for 5-7 days. Chronic stimulation induced a 50% reduction in evoked endplate potential (EPP) amplitude at stimulated neuromuscular junctions (NMJs). These changes appear to be presynaptic since miniature EPP (mEPP) amplitude was unchanged while mEPP frequency was decreased by 46% and paired-pulse facilitation was increased by 26%. High frequency facilitation (40 Hz, 2 s) was also increased by 89%. Moreover, stimulated NMJs presented a 92% decrease in synaptic depression (40 Hz, 2 s). An increase in mitochondrial metabolism was observed as indicated by a more pronounced labelling of active mitochondria (Mitotracker) in stimulated nerve terminals, which could account for their greater resistance to synaptic depression. NMJ length visualized by alpha-bungarotoxin staining of nAChRs was not affected. Presynaptic calcium signals measured with Calcium Green-1 were larger in stimulated NMJs at low frequency (0.2 Hz) and not different from control NMJs at higher frequency (40 Hz, 2 s and 30 s). These results suggest that some mechanisms downstream of calcium entry are responsible for the determination of synaptic output, such as a down-regulation of some calcium-binding proteins, which could explain the observed results. The possibility of a change in frequenin expression, a calcium-binding protein that is more prominently expressed in phasic synapses, was, however, refuted by our results.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources