

Evidence that entry into sporulation in Bacillus subtilis is governed by a gradual increase in the level and activity of the master regulator Spo0A
- PMID: 16166384
- PMCID: PMC1221893
- DOI: 10.1101/gad.1335705
Evidence that entry into sporulation in Bacillus subtilis is governed by a gradual increase in the level and activity of the master regulator Spo0A
Abstract
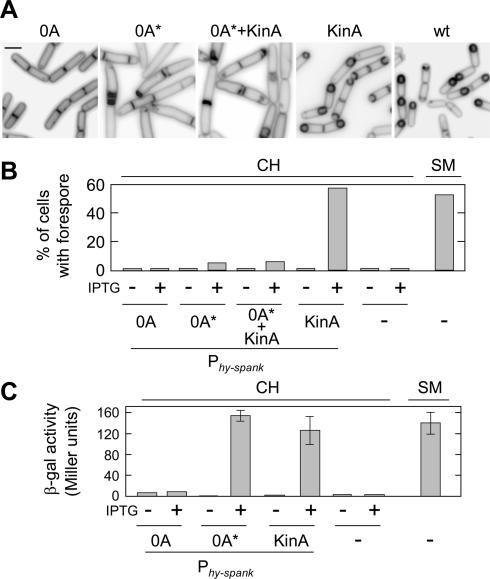
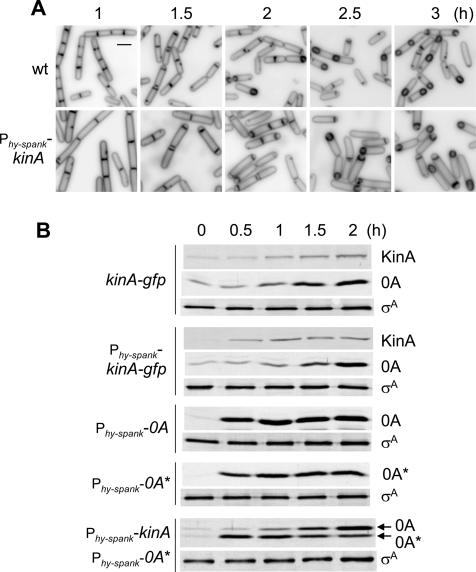
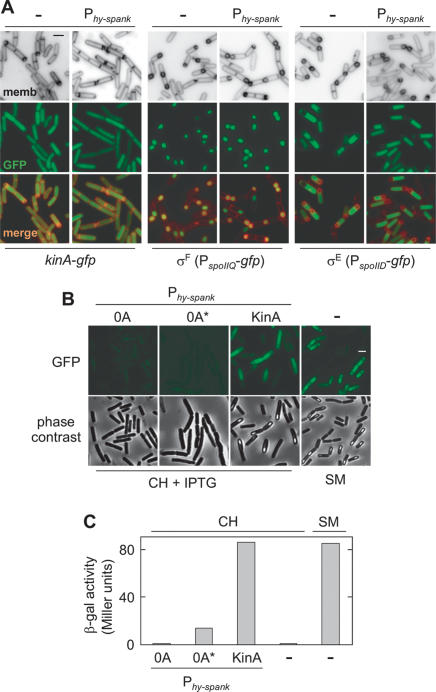
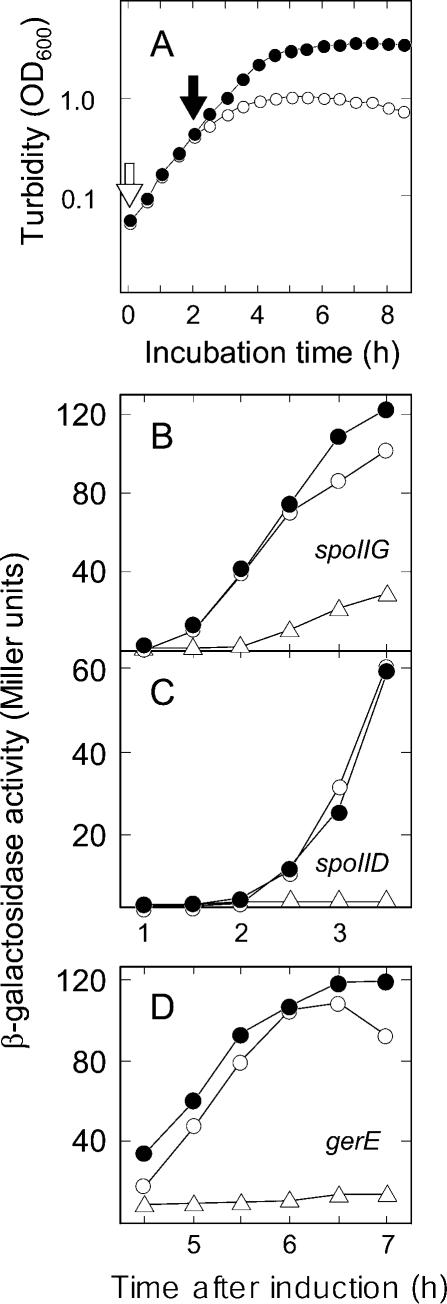
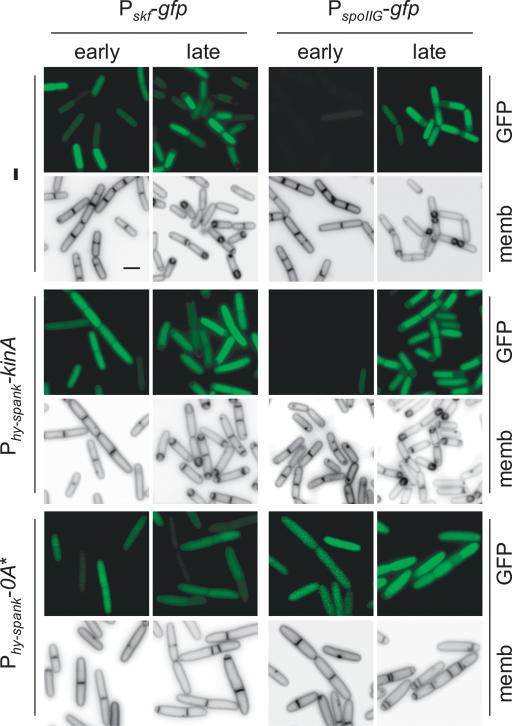
The transcription factor Spo0A is a master regulator for entry into sporulation in the bacterium Bacillus subtilis, but it has been uncertain whether activation of Spo0A is sufficient to trigger development. Spo0A, a member of the response regulator family of gene-control proteins, is activated by phosphorylation via a multicomponent phosphorelay in response to conditions of nutrient limitation. We now report that sporulation can be triggered with high efficiency in cells in the exponential phase of growth in rich medium by artificial induction of the synthesis of any one of three histidine kinases that feed phosphoryl groups into the relay. We further show that the levels of Spo0A protein and activity increase gradually over the first 2 h of sporulation both under conditions of nutrient limitation and in response to induction of kinase synthesis. Evidence indicates that this gradual increase in Spo0A protein and activity plays a critical role in triggering sporulation and requires the action of the phosphorelay.
Figures
References
-
- Ben-Yehuda S., Rudner, D.Z., and Losick, R. 2003. RacA, a bacterial protein that anchors chromosomes to the cell poles. Science 299: 532-536. - PubMed
-
- Burbulys D., Trach, K.A., and Hoch, J.A. 1991. Initiation of sporulation in B. subtilis is controlled by a multicomponent phosphorelay. Cell 64: 545-552. - PubMed
-
- Burkholder W.F., Kurtser, I., and Grossman, A.D. 2001. Replication initiation proteins regulate a developmental check-point in Bacillus subtilis. Cell 104: 269-279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases