

Imbalance between cysteine proteases and inhibitors in a baboon model of bronchopulmonary dysplasia
- PMID: 16166622
- PMCID: PMC2662933
- DOI: 10.1164/rccm.200503-425OC
Imbalance between cysteine proteases and inhibitors in a baboon model of bronchopulmonary dysplasia
Abstract
Rationale: Bronchopulmonary dysplasia (BPD) continues to be a major morbidity in preterm infants. The lung pathology in BPD is characterized by impaired alveolar and capillary development. An imbalance between proteases and protease inhibitors in association with changes in lung elastic fibers has been implicated in the pathogenesis of BPD.
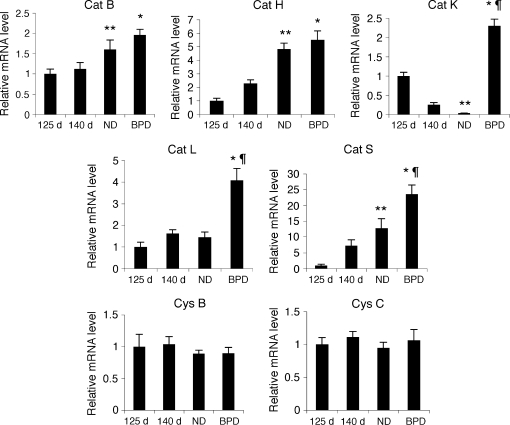
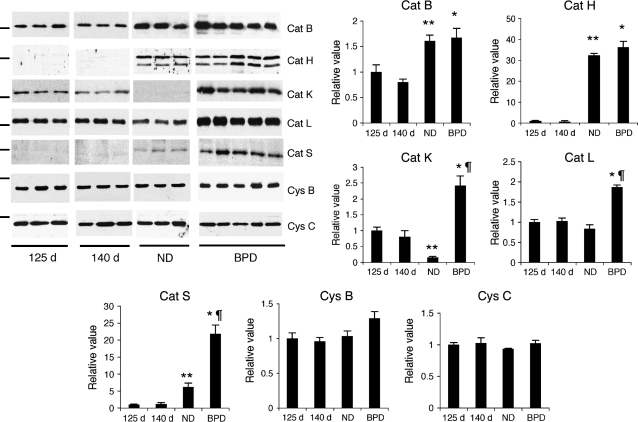
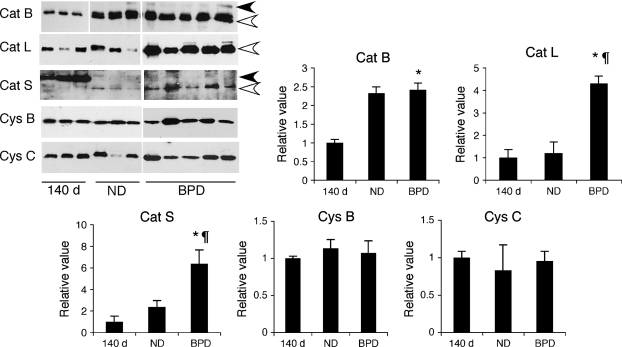
Objective: To investigate the expression and activity levels of papain-like lysosomal cysteine proteases, cathepsins B, H, K, L, S, and their inhibitors, cystatins B and C, in a baboon model of BPD.
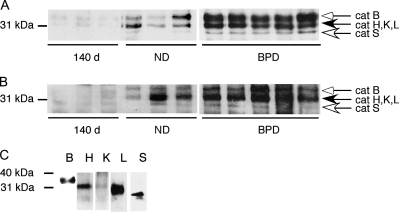
Methods: Real-time reverse transcriptase-polymerase chain reaction, immunohistochemistry, immunoblotting, active site labeling of cysteine proteases, and in situ hybridization were performed.
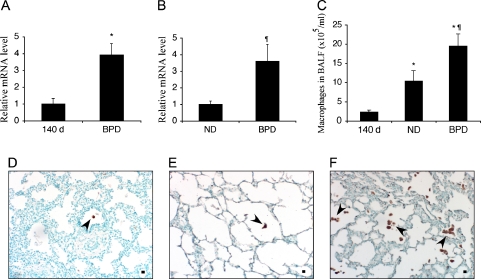
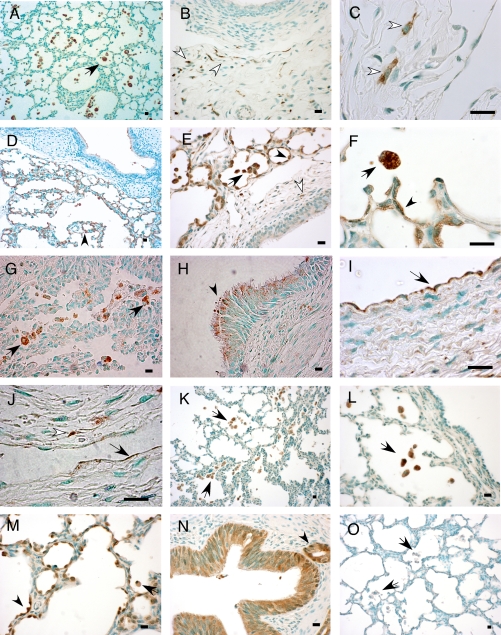
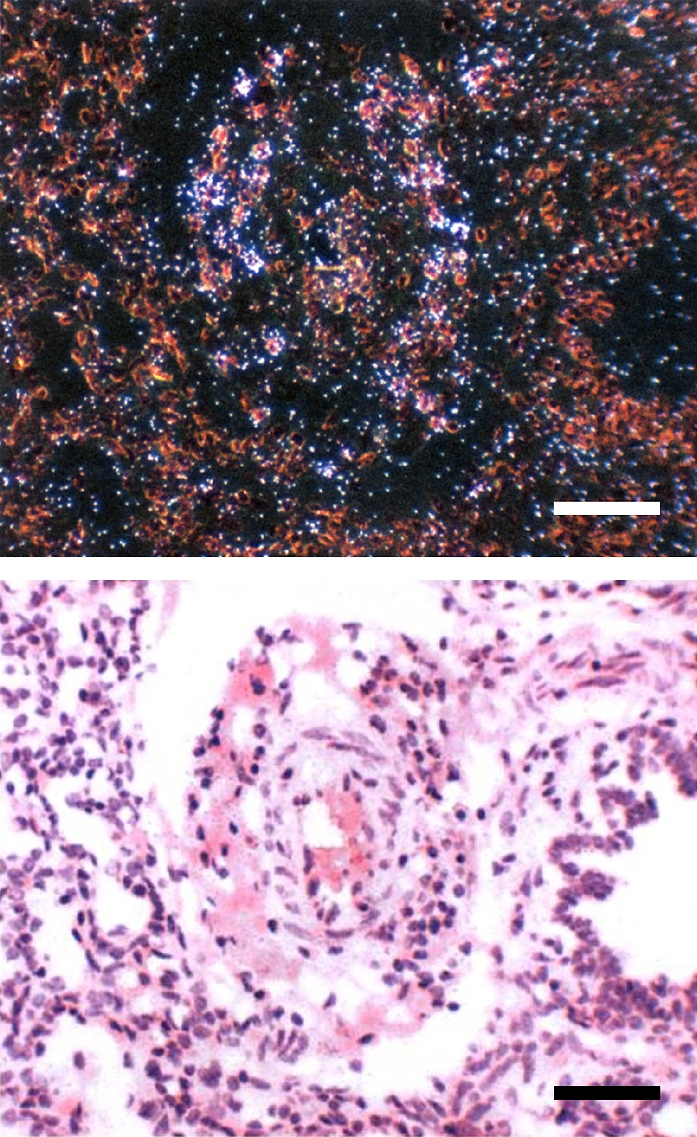
Measurements and main results: The steady-state mRNA and protein levels of all cathepsins were significantly increased in the lung tissue of baboons with BPD. In contrast, the steady-state mRNA and protein levels of two major cysteine protease inhibitors, cystatin B and C, were unchanged. Correlating with these alterations, the activity of cysteine proteases in lung tissue homogenates and bronchoalveolar lavage fluid was significantly higher in the BPD group. The levels of cathepsin B, H, and S increased and cathepsin K decreased with advancing gestation. All cathepsins, except for cat K, were immunolocalized to macrophages in BPD. In addition, cathepsin H and cystatin B were colocalized in type 2 alveolar epithelial cells. Cathepsin L was detected in some bronchial epithelial, endothelial, and interstitial cells. Cathepsin K was localized to some perivascular cells by in situ hybridization.
Conclusions: Cumulatively, these findings demonstrate an imbalance between cysteine proteases and their inhibitors in BPD.
Figures
References
-
- Northway WJ, Rosan RC, Porter DY. Pulmonary disease following respirator therapy of hyaline-membrane disease: bronchopulmonary dysplasia. N Engl J Med 1967;276:357–368. - PubMed
-
- Bonikos DS, Bensch KG. Pathogenesis of bronchopulmonary dysplasia. In: Merritt TA, Northway WH, editors. Bronchopulmonary dysplasia. Boston, MA: Blackwell; 1988. pp. 33–58.
-
- Bancalari E, Claure N, Sosenko IR. Bronchopulmonary dysplasia: changes in pathogenesis, epidemiology and definition. Semin Neonatol 2003;8:63–71. - PubMed
-
- Tanswell AK, Jankov RP. Bronchopulmonary dysplasia: one disease or two? Am J Respir Crit Care Med 2003;167:1–2. - PubMed
-
- Coalson JJ. Pathology of new bronchopulmonary dysplasia. Semin Neonatol 2003;8:73–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
