

HATs and HDACs in neurodegeneration: a tale of disconcerted acetylation homeostasis
- PMID: 16167067
- PMCID: PMC1963416
- DOI: 10.1038/sj.cdd.4401769
HATs and HDACs in neurodegeneration: a tale of disconcerted acetylation homeostasis
Abstract
Gradual disclosure of the molecular basis of selective neuronal apoptosis during neurodegenerative diseases reveals active participation of acetylating and deacetylating agents during the process. Several studies have now successfully manipulated neuronal vulnerability by influencing the dose and enzymatic activity of histone acetyltransferases (HATs) and histone deacetylases (HDACs), enzymes regulating acetylation homeostasis within the nucleus, thus focusing on the importance of balanced acetylation status in neuronal vitality. It is now increasingly becoming clear that acetylation balance is greatly impaired during neurodegenerative conditions. Herein, we attempt to illuminate molecular means by which such impairment is manifested and how the compromised acetylation homeostasis is intimately coupled to neurodegeneration. Finally, we discuss the therapeutic potential of reinstating the HAT-HDAC balance to ameliorate neurodegenerative diseases.
Figures
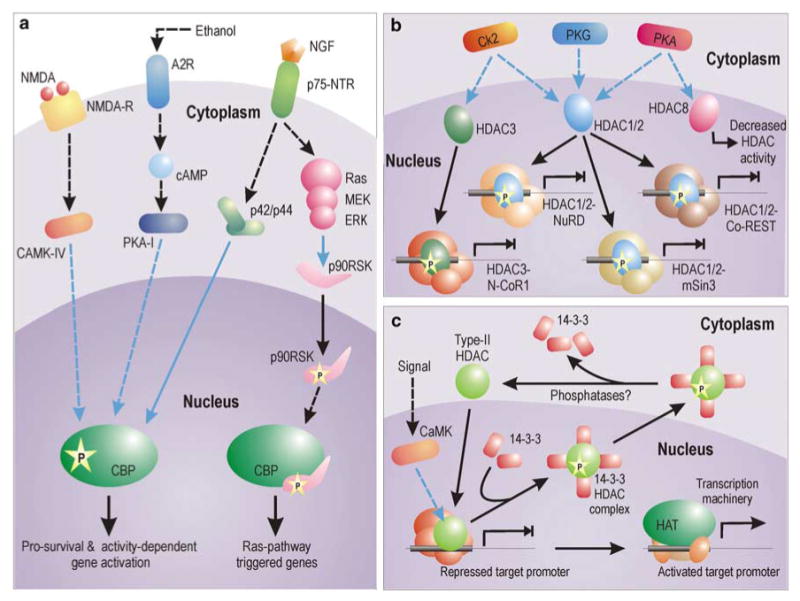
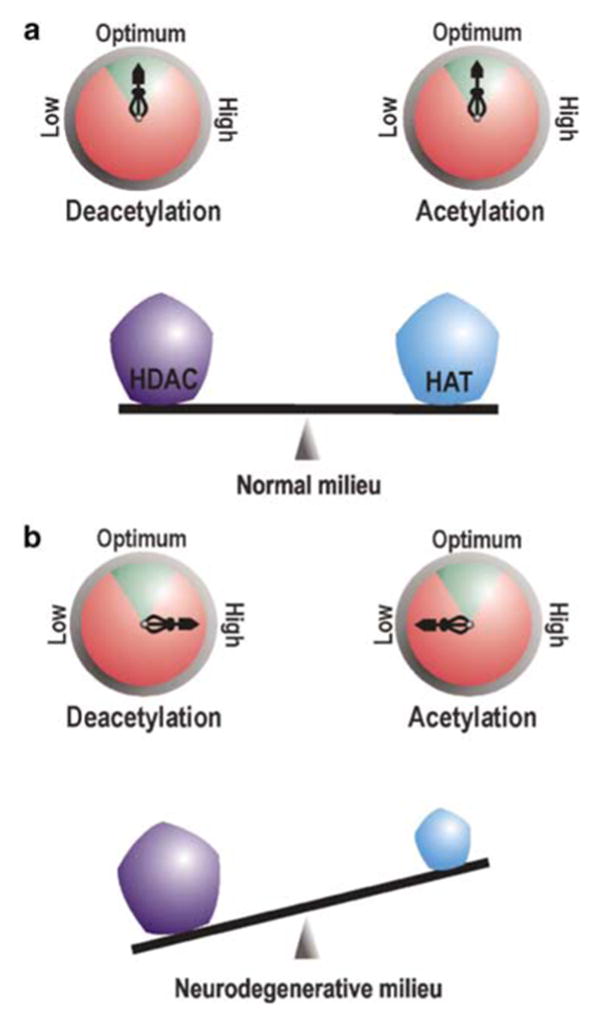
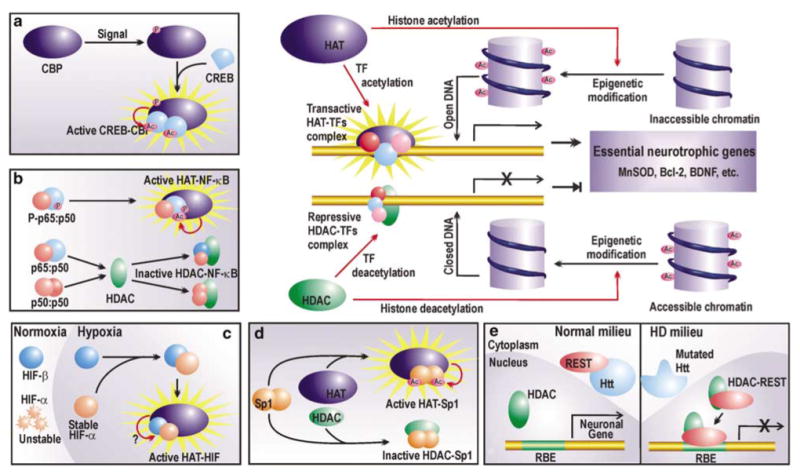
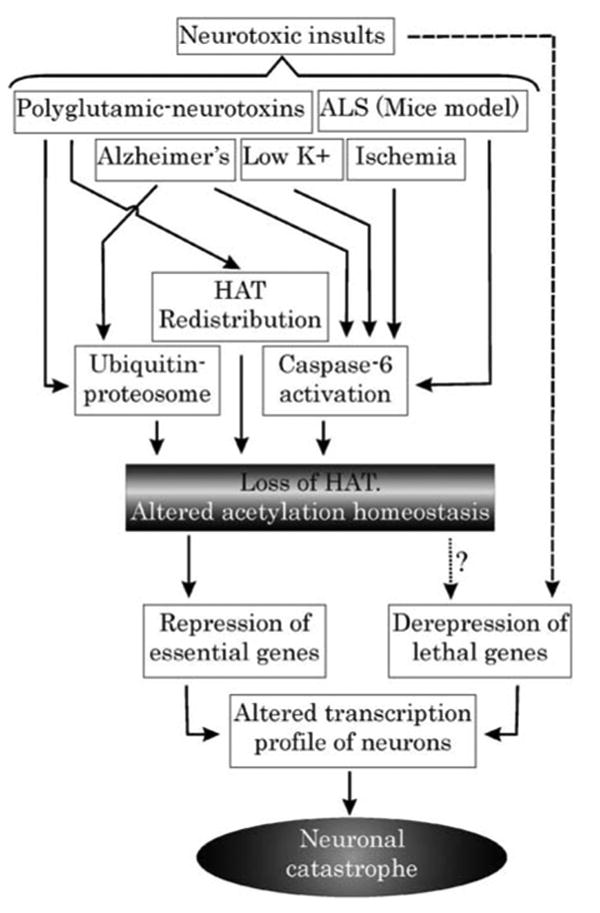
References
-
- Taunton J, Hassig CA, Schreiber SL. A mammalian histone deacetylase related to the yeast transcriptional regulator Rpd3p. Science. 1996;272:408–411. - PubMed
-
- Marmorstein R, Roth SY. Histone acetyltransferases: function, structure, and catalysis. Curr Opin Genet Dev. 2001;11:155–161. - PubMed
-
- Thiagalingam S, Cheng KH, Lee HJ, Mineva N, Thiagalingam A, Ponte JF. Histone deacetylases: unique players in shaping the epigenetic histone code. Ann N Y Acad Sci. 2003;983:84–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
