

Amyloid fibril formation can proceed from different conformations of a partially unfolded protein
- PMID: 16169975
- PMCID: PMC1366985
- DOI: 10.1529/biophysj.105.068726
Amyloid fibril formation can proceed from different conformations of a partially unfolded protein
Abstract
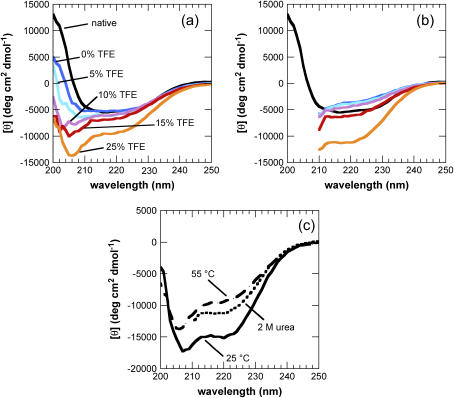
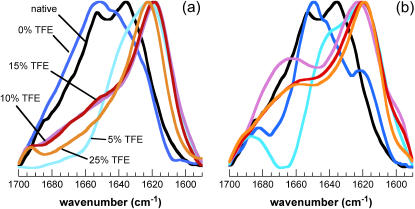
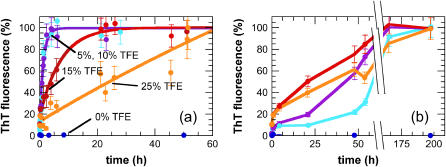
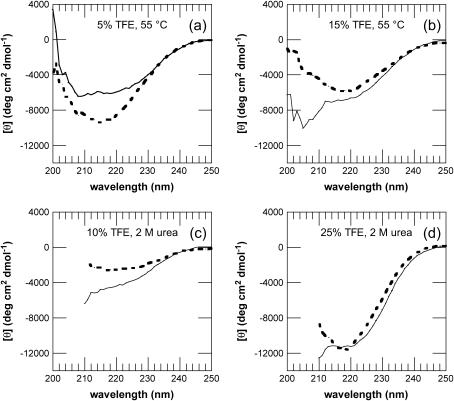
Protein misfolding and aggregation are interconnected processes involved in a wide variety of nonneuropathic, systemic, and neurodegenerative diseases. More generally, if mutations in sequence or changes in environmental conditions lead to partial unfolding of the native state of a protein, it will often aggregate, sometimes into well-defined fibrillar structures. A great deal of interest has been directed at discovering the characteristic features of metastable partially unfolded states that precede the aggregated states of proteins. In this work, human muscle acylphosphatase (AcP) has been first destabilized, by addition of urea or by means of elevated temperatures, and then incubated in the presence of different concentrations of 2,2,2, trifluoroethanol ranging from 5% to 25% (v/v). The results show that AcP is able to form both fibrillar and nonfibrillar aggregates with a high beta-sheet content from partially unfolded states with very different structural features. Moreover, the presence of alpha-helical structure in such a state does not appear to be a fundamental determinant of the ability to aggregate. The lack of ready aggregation under some of the conditions examined here is attributable primarily to the intrinsic properties of the solutions rather than to specific structural features of the partially unfolded states that precede aggregation. Aggregation appears to be favored when the solution conditions promote stable intermolecular interactions, particularly hydrogen bonds. In addition, the structures of the resulting aggregates are largely independent of the conformational properties of their soluble precursors.
Figures
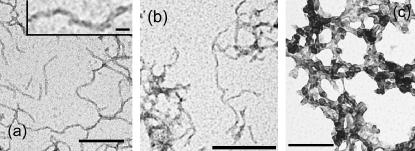
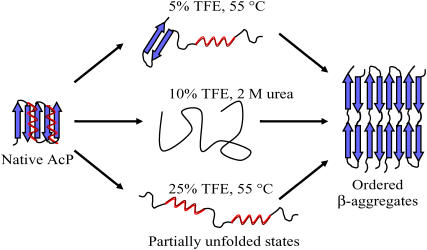
References
-
- Dobson, C. M. 2003. Protein folding and misfolding. Nature. 426:884–890. - PubMed
-
- Walsh, D. M., and D. J. Selkoe. 2004. Oligomers on the brain: the emerging role of soluble protein aggregates in neurodegeneration. Protein Pept. Lett. 11:213–228. - PubMed
-
- Buxbaum, J. N. 2004. The systemic amyloidoses. Curr. Opin. Rheumatol. 16:67–75. - PubMed
-
- Stefani, M., and C. M. Dobson. 2003. Protein aggregation and aggregate toxicity: new insights into protein folding, misfolding diseases and biological evolution. J. Mol. Med. 81:678–699. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
