

The structure of human CD23 and its interactions with IgE and CD21
- PMID: 16172256
- PMCID: PMC2212946
- DOI: 10.1084/jem.20050811
The structure of human CD23 and its interactions with IgE and CD21
Abstract
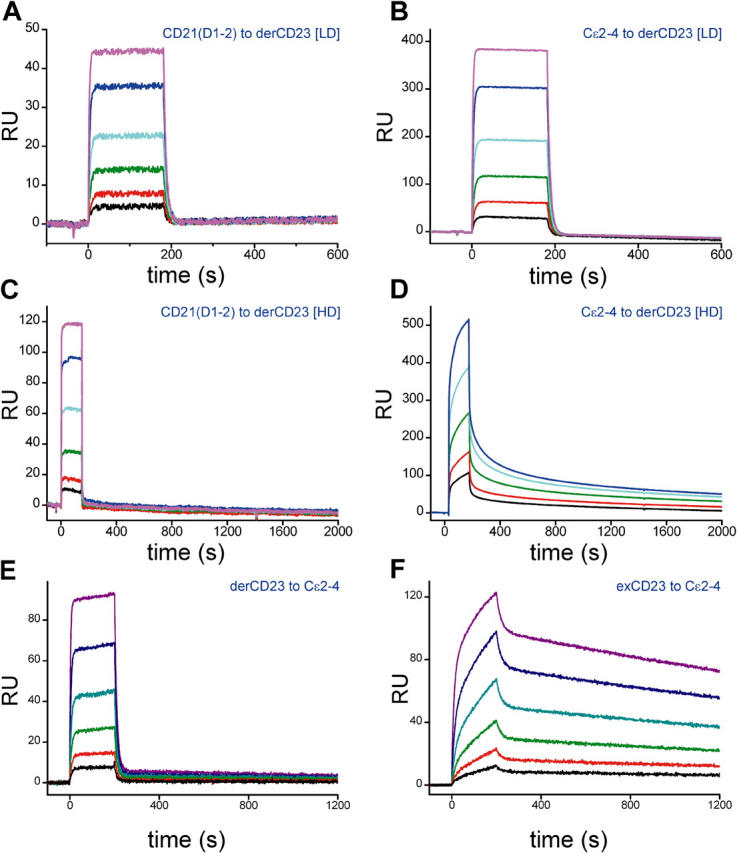
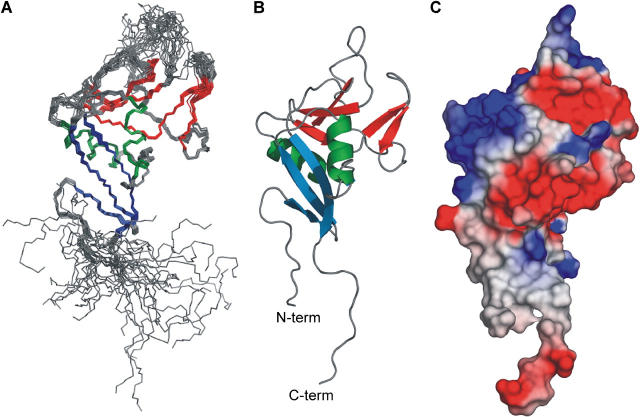
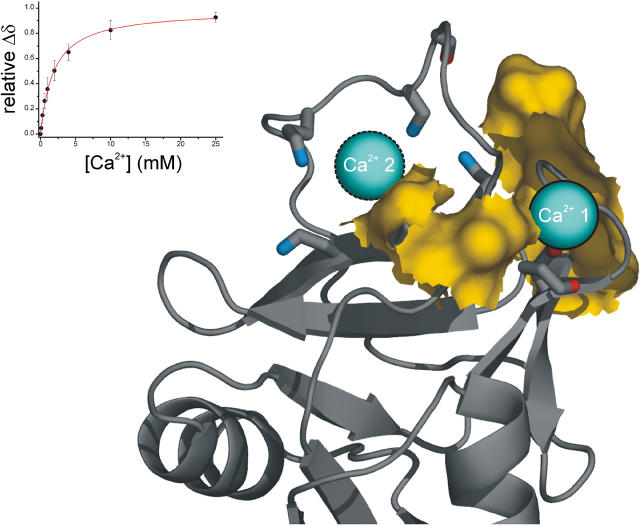
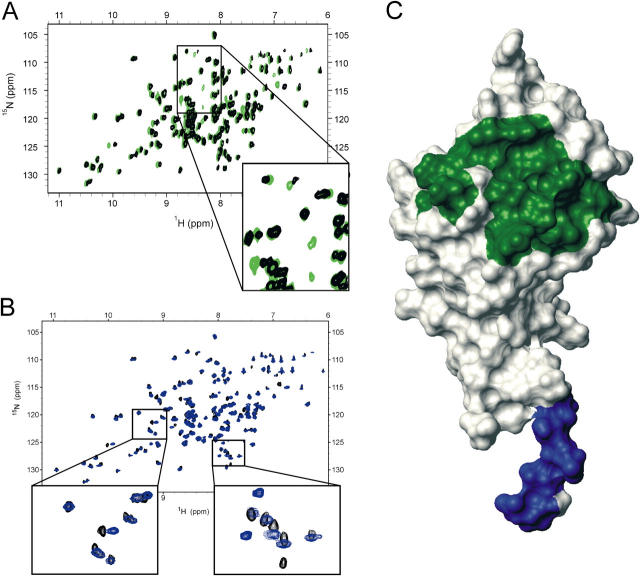
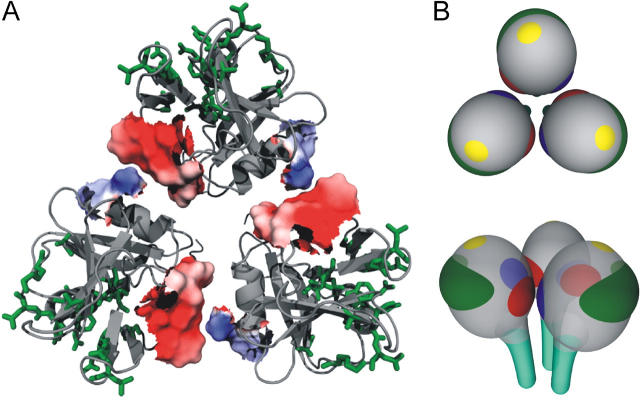
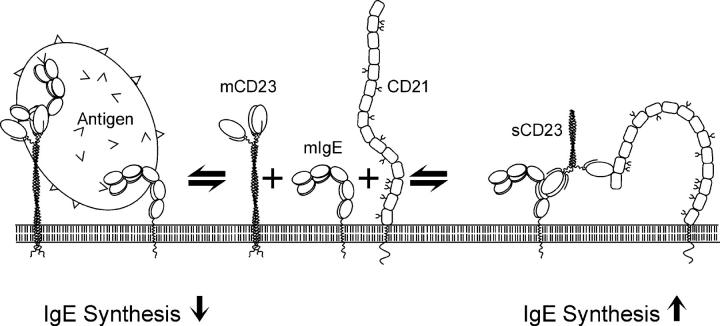
The low-affinity immunoglobulin E (IgE) receptor, CD23 (FcepsilonRII), binds both IgE and CD21 and, through these interactions, regulates the synthesis of IgE, the antibody isotype that mediates the allergic response. We have determined the three-dimensional structure of the C-type lectin domain of CD23 in solution by nuclear magnetic resonance spectroscopy. An analysis of concentration-dependent chemical shift perturbations have allowed us to identify the residues engaged in self-association to the trimeric state, whereas ligand-induced changes have defined the binding sites for IgE and CD21. The results further reveal that CD23 can bind both ligands simultaneously. Despite the C-type lectin domain structure, none of the interactions require calcium. We also find that IgE and CD23 can interact to form high molecular mass multimeric complexes. The interactions that we have described provide a solution to the paradox that CD23 is involved in both up- and down-regulation of IgE and provide a structural basis for the development of inhibitors of allergic disease.
Figures
References
-
- Gould, H.J., R.L. Beavil, R. Reljic, J. Shi, C.W. Ma, B.J. Sutton, and R. Ghirlando. 1997. IgE homeostasis: is CD23 the safety switch? IgE Regulation: Molecular Mechanisms. D. Vercelli, editor. John Wiley & Sons Inc., New York. 35–59.
-
- Aubry, J.P., S. Pochon, P. Graber, K.U. Jansen, and J.Y. Bonnefoy. 1992. CD21 is a ligand for CD23 and regulates IgE production. Nature. 358:505–507. - PubMed
-
- Soilleux, E.J., R. Barten, and J. Trowsdale. 2000. DC-SIGN; a related gene, DC-SIGNR; and CD23 form a cluster on 19p13. J. Immunol. 165:2937–2942. - PubMed
-
- Shi, J., R. Ghirlando, R.L. Beavil, A.J. Beavil, M.B. Keown, R.J. Young, R.J. Owens, B.J. Sutton, and H.J. Gould. 1997. Interaction of the low-affinity receptor CD23/Fc epsilonRII lectin domain with the Fc epsilon3-4 fragment of human immunoglobulin E. Biochemistry. 36:2112–2122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
