

Molecular characterization of the major wheat domestication gene Q
- PMID: 16172507
- PMCID: PMC1456182
- DOI: 10.1534/genetics.105.044727
Molecular characterization of the major wheat domestication gene Q
Abstract
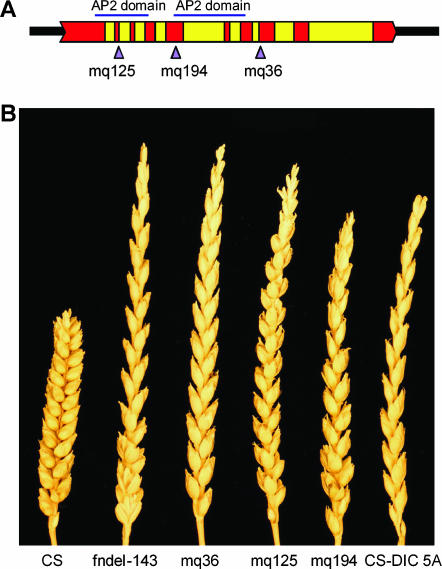
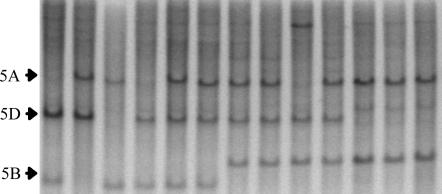
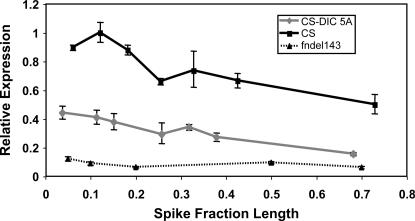
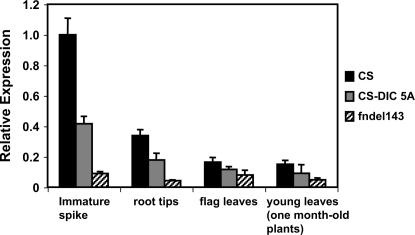
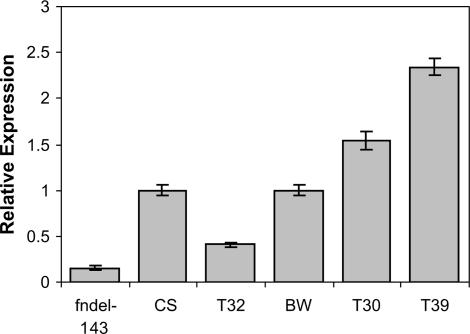
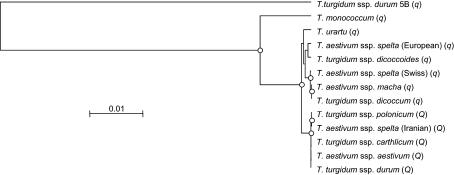
The Q gene is largely responsible for the widespread cultivation of wheat because it confers the free-threshing character. It also pleiotropically influences many other domestication-related traits such as glume shape and tenacity, rachis fragility, spike length, plant height, and spike emergence time. We isolated the Q gene and verified its identity by analysis of knockout mutants and transformation. The Q gene has a high degree of similarity to members of the AP2 family of transcription factors. The Q allele is more abundantly transcribed than q, and the two alleles differ for a single amino acid. An isoleucine at position 329 in the Q protein leads to an abundance of homodimer formation in yeast cells, whereas a valine in the q protein appears to limit homodimer formation. Ectopic expression analysis allowed us to observe both silencing and overexpression effects of Q. Rachis fragility, glume shape, and glume tenacity mimicked the q phenotype in transgenic plants exhibiting post-transcriptional silencing of the transgene and the endogenous Q gene. Variation in spike compactness and plant height were associated with the level of transgene transcription due to the dosage effects of Q. The q allele is the more primitive, and the mutation that gave rise to Q occurred only once leading to the world's cultivated wheats.
Figures



References
-
- Altpeter, F., V. Vasil, V. Srivastava and I. K. Vasil, 1996. Integration and expression of the high-molecular-weight glutenin subunit 1Ax1 gene into wheat. Nat. Biotech. 14: 1155–1159. - PubMed
-
- Bowman, J. L., J. Alvarez, D. Weigel, E. M. Meyerowitz and D. R. Smyth, 1993. Control of flower development in Arabidopsis thaliana by APETALA1 and interacting genes. Development 119: 721–743.
-
- Cenci, A., N. Chantret, X. Kong, Y. Gu, O. D. Anderson et al., 2003. Construction and characterization of a half million clone BAC library of durum wheat (Triticum turgidum ssp. durum). Theor. Appl. Genet. 107: 931–939. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
