RNA catalysis in model protocell vesicles
- PMID: 16173749
- PMCID: PMC5072289
- DOI: 10.1021/ja051784p
RNA catalysis in model protocell vesicles
Abstract
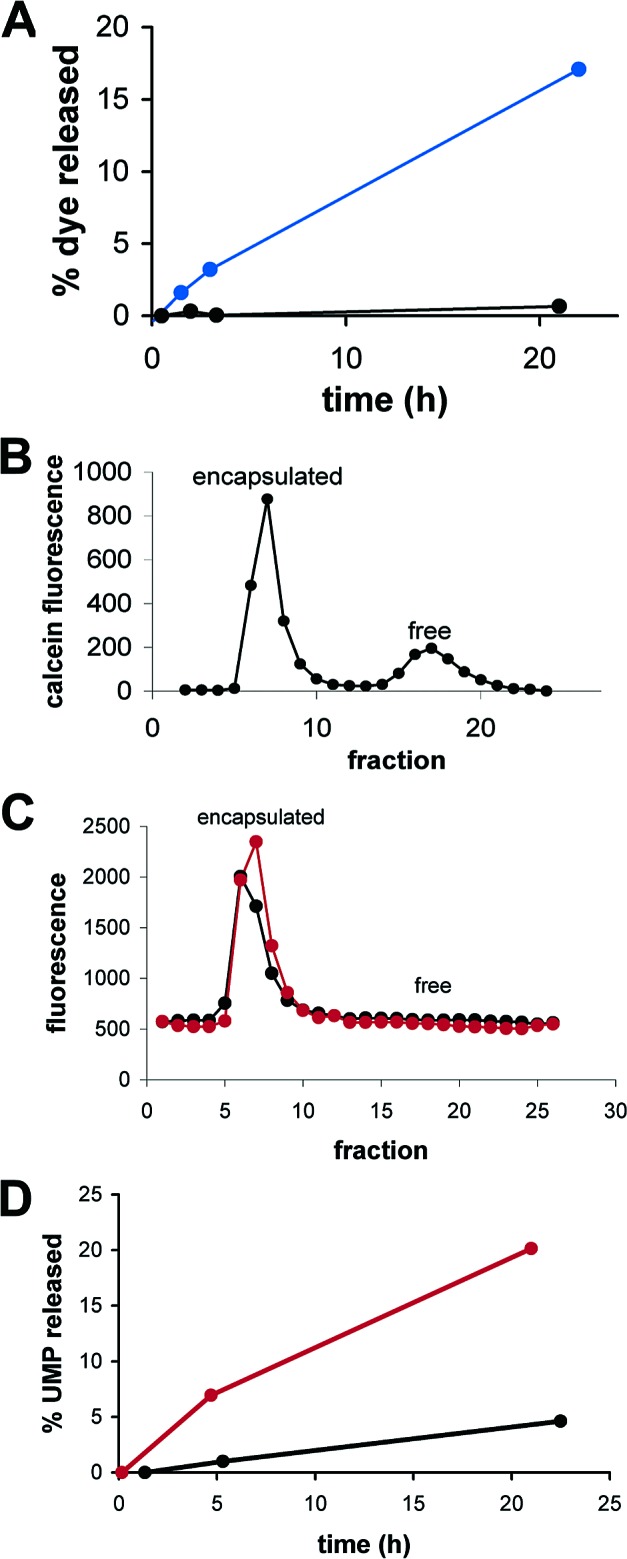
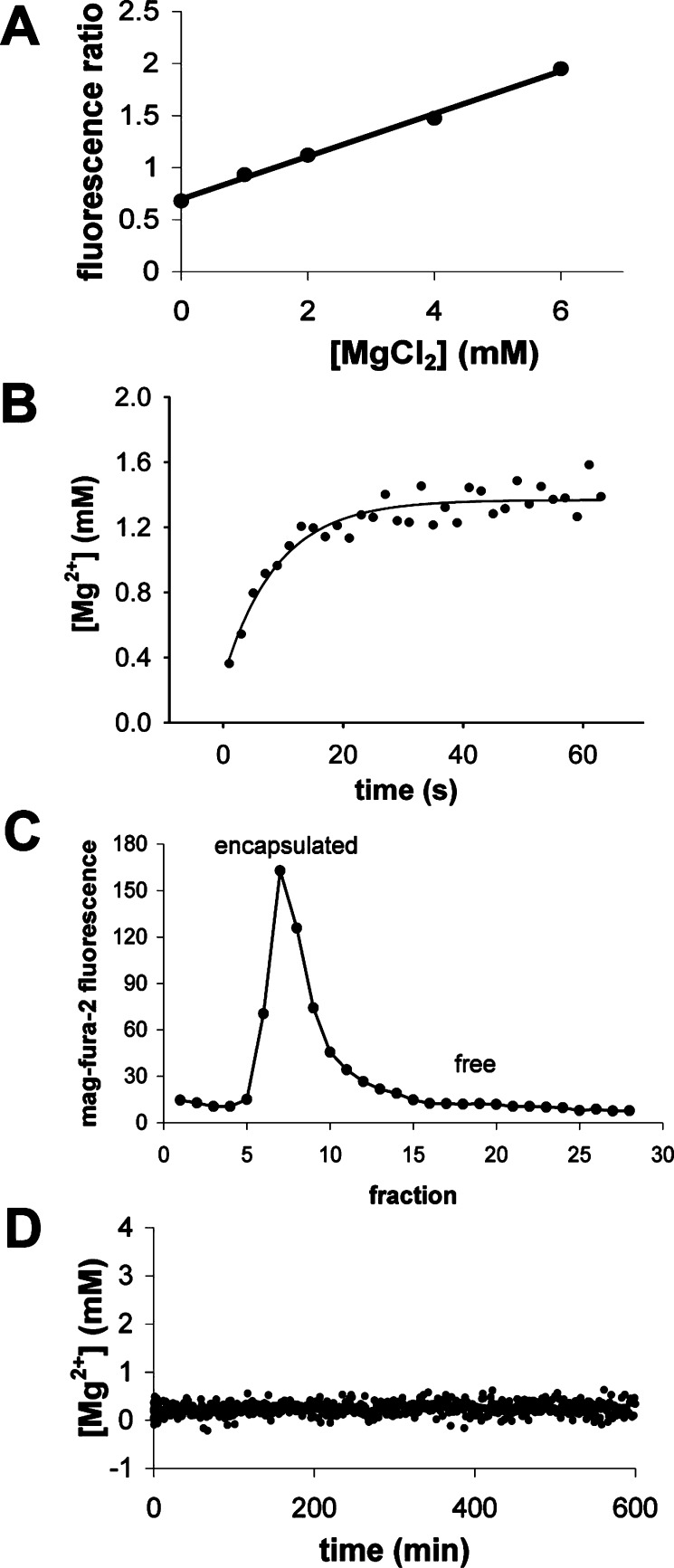
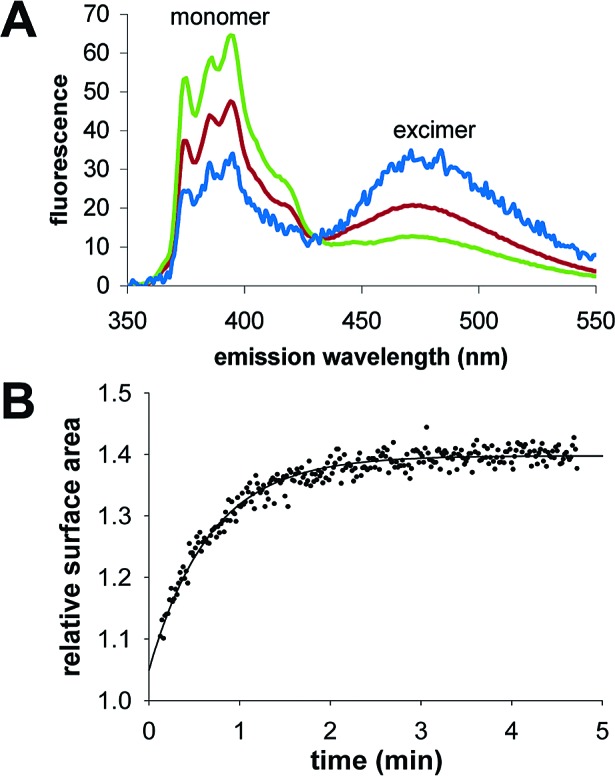
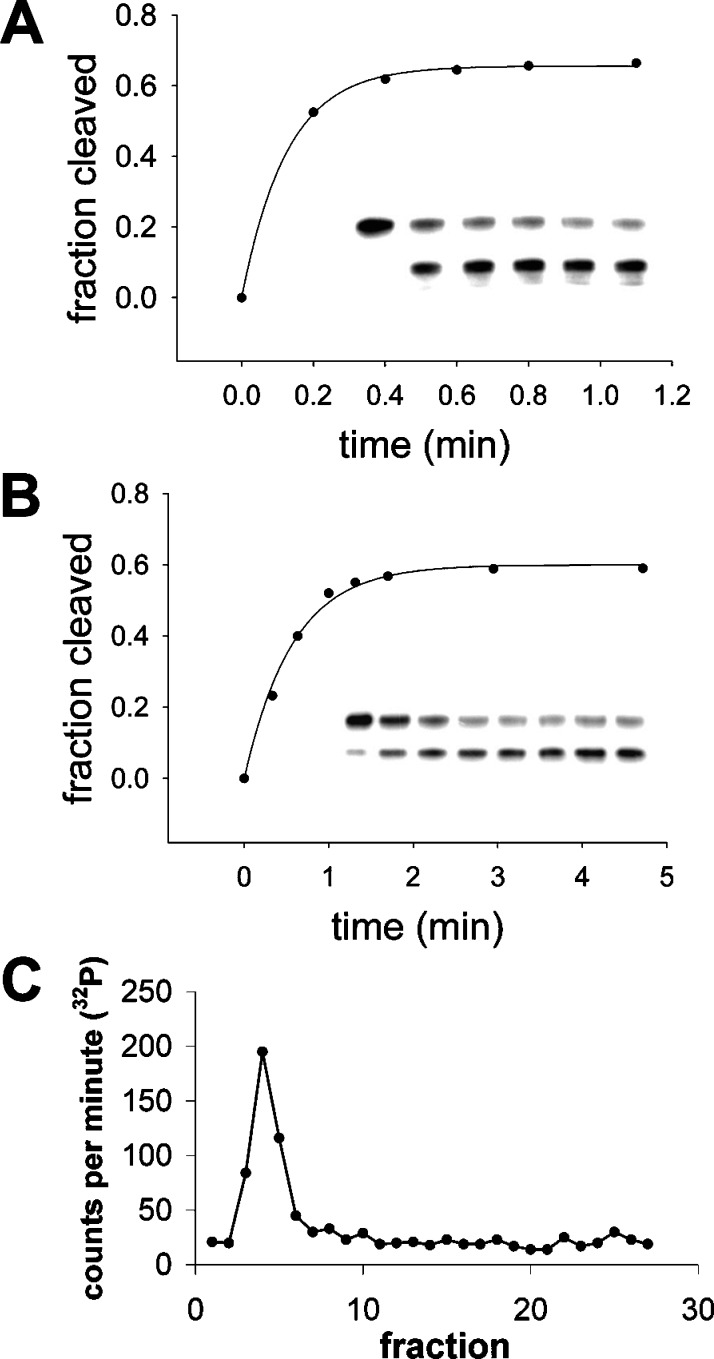
We are engaged in a long-term effort to synthesize chemical systems capable of Darwinian evolution, based on the encapsulation of self-replicating nucleic acids in self-replicating membrane vesicles. Here, we address the issue of the compatibility of these two replicating systems. Fatty acids form vesicles that are able to grow and divide, but vesicles composed solely of fatty acids are incompatible with the folding and activity of most ribozymes, because low concentrations of divalent cations (e.g., Mg(2+)) cause fatty acids to precipitate. Furthermore, vesicles that grow and divide must be permeable to the cations and substrates required for internal metabolism. We used a mixture of myristoleic acid and its glycerol monoester to construct vesicles that were Mg(2+)-tolerant and found that Mg(2+) cations can permeate the membrane and equilibrate within a few minutes. In vesicles encapsulating a hammerhead ribozyme, the addition of external Mg(2+) led to the activation and self-cleavage of the ribozyme molecules. Vesicles composed of these amphiphiles grew spontaneously through osmotically driven competition between vesicles, and further modification of the membrane composition allowed growth following mixed micelle addition. Our results show that membranes made from simple amphiphiles can form vesicles that are stable enough to retain encapsulated RNAs in the presence of divalent cations, yet dynamic enough to grow spontaneously and allow the passage of Mg(2+) and mononucleotides without specific macromolecular transporters. This combination of stability and dynamics is critical for building model protocells in the laboratory and may have been important for early cellular evolution.
Figures