

Two-component signal transduction pathways regulating growth and cell cycle progression in a bacterium: a system-level analysis
- PMID: 16176121
- PMCID: PMC1233412
- DOI: 10.1371/journal.pbio.0030334
Two-component signal transduction pathways regulating growth and cell cycle progression in a bacterium: a system-level analysis
Abstract
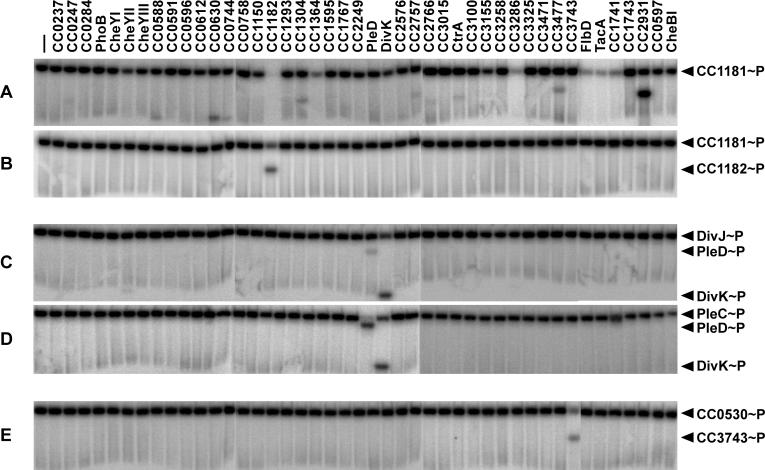
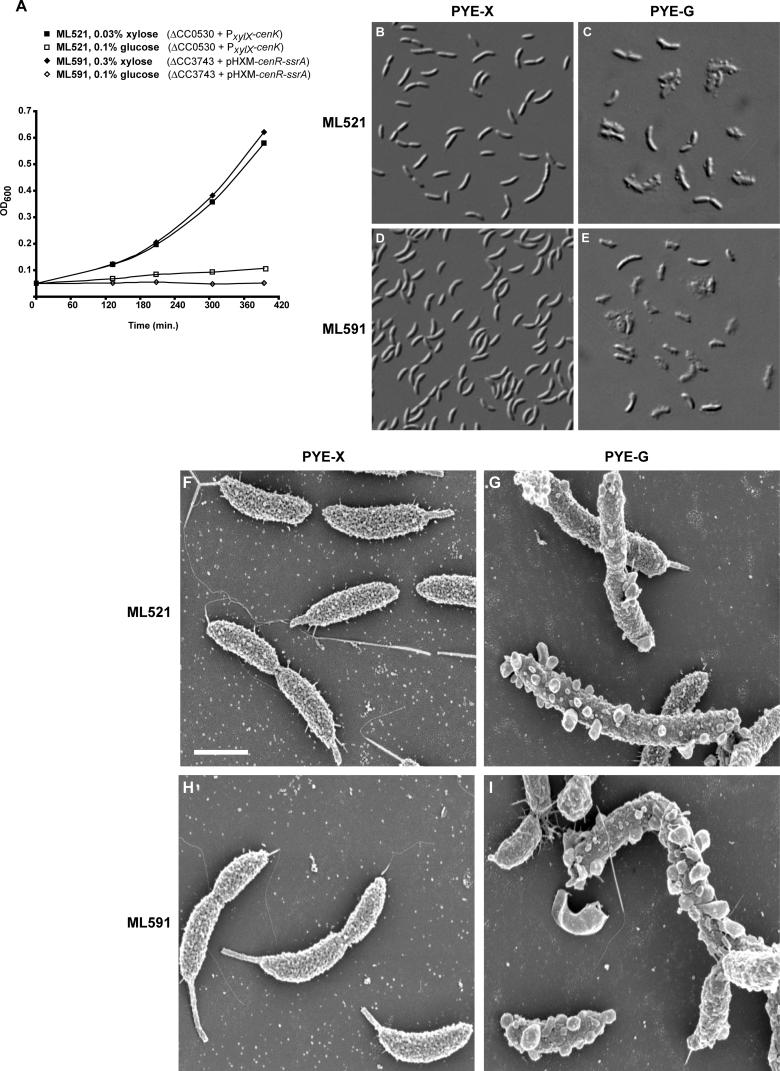
Two-component signal transduction systems, comprised of histidine kinases and their response regulator substrates, are the predominant means by which bacteria sense and respond to extracellular signals. These systems allow cells to adapt to prevailing conditions by modifying cellular physiology, including initiating programs of gene expression, catalyzing reactions, or modifying protein-protein interactions. These signaling pathways have also been demonstrated to play a role in coordinating bacterial cell cycle progression and development. Here we report a system-level investigation of two-component pathways in the model organism Caulobacter crescentus. First, by a comprehensive deletion analysis we show that at least 39 of the 106 two-component genes are required for cell cycle progression, growth, or morphogenesis. These include nine genes essential for growth or viability of the organism. We then use a systematic biochemical approach, called phosphotransfer profiling, to map the connectivity of histidine kinases and response regulators. Combining these genetic and biochemical approaches, we identify a new, highly conserved essential signaling pathway from the histidine kinase CenK to the response regulator CenR, which plays a critical role in controlling cell envelope biogenesis and structure. Depletion of either cenK or cenR leads to an unusual, severe blebbing of cell envelope material, whereas constitutive activation of the pathway compromises cell envelope integrity, resulting in cell lysis and death. We propose that the CenK-CenR pathway may be a suitable target for new antibiotic development, given previous successes in targeting the bacterial cell wall. Finally, the ability of our in vitro phosphotransfer profiling method to identify signaling pathways that operate in vivo takes advantage of an observation that histidine kinases are endowed with a global kinetic preference for their cognate response regulators. We propose that this system-wide selectivity insulates two-component pathways from one another, preventing unwanted cross-talk.
Figures





References
-
- Ryan KR, Shapiro L. Temporal and spatial regulation in prokaryotic cell cycle progression and development. Annu Rev Biochem. 2003;72:367–394. - PubMed
-
- McAdams HH, Shapiro L. A bacterial cell-cycle regulatory network operating in time and space. Science. 2003;301:1874–1877. - PubMed
-
- Skerker JM, Laub MT. Cell-cycle progression and the generation of asymmetry in Caulobacter crescentus . Nat Rev Microbiol. 2004;2:325–337. - PubMed
-
- Stock AM, Robinson VL, Goudreau PN. Two-component signal transduction. Annu Rev Biochem. 2000;69:183–215. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
