

Identification of a novel sialic acid transporter in Haemophilus ducreyi
- PMID: 16177350
- PMCID: PMC1230923
- DOI: 10.1128/IAI.73.10.6727-6735.2005
Identification of a novel sialic acid transporter in Haemophilus ducreyi
Abstract
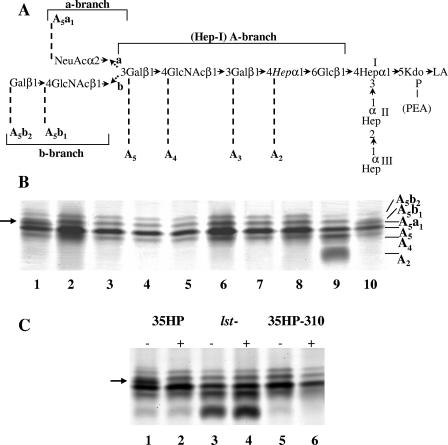
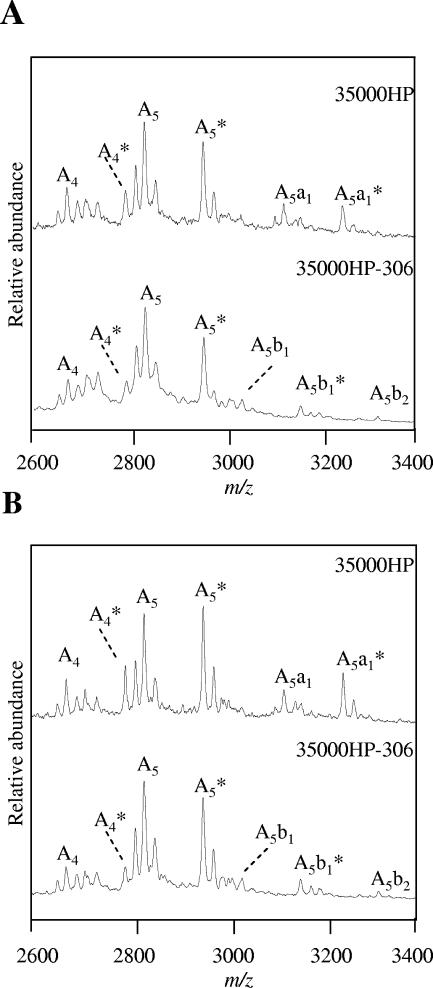
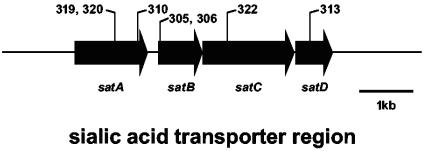
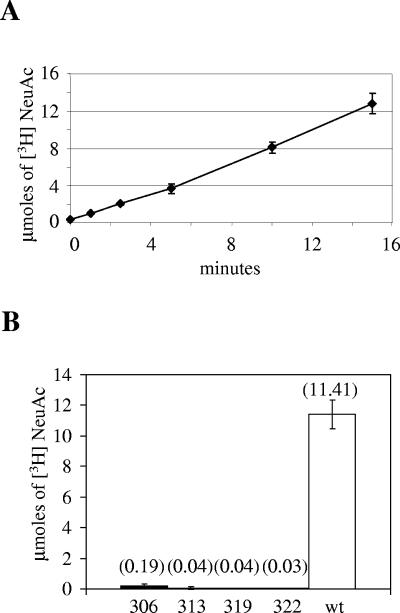
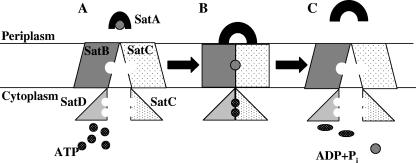
Haemophilus ducreyi, the causative agent of chancroid, produces a lipooligosaccharide (LOS) which terminates in N-acetyllactosamine. This glycoform can be further extended by the addition of a single sialic acid residue to the terminal galactose moiety. H. ducreyi does not synthesize sialic acid, which must be acquired from the host during infection or from the culture medium when the bacteria are grown in vitro. However, H. ducreyi does not have genes that are highly homologous to the genes encoding known bacterial sialic acid transporters. In this study, we identified the sialic acid transporter by screening strains in a library of random transposon mutants for those mutants that were unable to add sialic acid to N-acetyllactosamine-containing LOS. Mutants that reacted with the monoclonal antibody 3F11, which recognizes the terminal lactosamine structure, and lacked reactivity with the lectin Maackia amurensis agglutinin, which recognizes alpha2,3-linked sialic acid, were further characterized to demonstrate that they produced a N-acetyllactosamine-containing LOS by silver-stained sodium dodecyl sulfate-polyacrylamide gel electrophoresis and mass spectrometric analyses. The genes interrupted in these mutants were mapped to a four-gene cluster with similarity to genes encoding bacterial ABC transporters. Uptake assays using radiolabeled sialic acid confirmed that the mutants were unable to transport sialic acid. This study is the first report of bacteria using an ABC transporter for sialic acid uptake.
Figures
Similar articles
-
Haemophilus ducreyi produces a novel sialyltransferase. Identification of the sialyltransferase gene and construction of mutants deficient in the production of the sialic acid-containing glycoform of the lipooligosaccharide.J Biol Chem. 1999 Feb 12;274(7):4106-14. doi: 10.1074/jbc.274.7.4106. J Biol Chem. 1999. PMID: 9933604
-
Characterization of a transposon Tn916-generated mutant of Haemophilus ducreyi 35000 defective in lipooligosaccharide biosynthesis.J Bacteriol. 1997 Aug;179(16):5062-71. doi: 10.1128/jb.179.16.5062-5071.1997. J Bacteriol. 1997. PMID: 9260947 Free PMC article.
-
Construction and characterization of Haemophilus ducreyi lipooligosaccharide (LOS) mutants defective in expression of heptosyltransferase III and beta1,4-glucosyltransferase: identification of LOS glycoforms containing lactosamine repeats.Infect Immun. 2000 Jun;68(6):3352-61. doi: 10.1128/IAI.68.6.3352-3361.2000. Infect Immun. 2000. PMID: 10816485 Free PMC article.
-
Bacterial Sialic Acid Catabolism at the Host-Microbe Interface.J Microbiol. 2023 Apr;61(4):369-377. doi: 10.1007/s12275-023-00035-7. Epub 2023 Mar 27. J Microbiol. 2023. PMID: 36972004 Review.
-
Free sialic acid storage disorder: Progress and promise.Neurosci Lett. 2021 Jun 11;755:135896. doi: 10.1016/j.neulet.2021.135896. Epub 2021 Apr 20. Neurosci Lett. 2021. PMID: 33862140 Free PMC article. Review.
Cited by
-
Identification of Haemophilus ducreyi genes expressed during human infection.Microbiology (Reading). 2008 Apr;154(Pt 4):1152-1160. doi: 10.1099/mic.0.2007/013953-0. Microbiology (Reading). 2008. PMID: 18375807 Free PMC article.
-
Molecular characterization of the interaction of sialic acid with the periplasmic binding protein from Haemophilus ducreyi.J Biol Chem. 2018 Dec 28;293(52):20073-20084. doi: 10.1074/jbc.RA118.005151. Epub 2018 Oct 12. J Biol Chem. 2018. PMID: 30315109 Free PMC article.
-
An infant-associated bacterial commensal utilizes breast milk sialyloligosaccharides.J Biol Chem. 2011 Apr 8;286(14):11909-18. doi: 10.1074/jbc.M110.193359. Epub 2011 Feb 2. J Biol Chem. 2011. PMID: 21288901 Free PMC article.
-
The Sodium Sialic Acid Symporter From Staphylococcus aureus Has Altered Substrate Specificity.Front Chem. 2018 Jul 4;6:233. doi: 10.3389/fchem.2018.00233. eCollection 2018. Front Chem. 2018. PMID: 30023356 Free PMC article.
-
Sialic acid catabolism confers a competitive advantage to pathogenic vibrio cholerae in the mouse intestine.Infect Immun. 2009 Sep;77(9):3807-16. doi: 10.1128/IAI.00279-09. Epub 2009 Jun 29. Infect Immun. 2009. PMID: 19564383 Free PMC article.
References
-
- Alfa, M. J., and P. DeGagne. 1997. Attachment of Haemophilus ducreyi to human foreskin fibroblasts involves LOS and fibronectin. Microb. Pathog. 22:39-46. - PubMed
-
- Apicella, M. A., R. E. Mandrell, M. Shero, M. E. Wilson, J. M. Griffiss, G. F. Brooks, C. Lammel, J. F. Breen, and P. A. Rice. 1990. Modification by sialic acid of Neisseria gonorrhoeae lipooligosaccharide epitope expression in human urethral exudates: an immunoelectron microscopic analysis. J. Infect. Dis. 162:506-512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
