

How to activate a plant gravireceptor. Early mechanisms of gravity sensing studied in characean rhizoids during parabolic flights
- PMID: 16183834
- PMCID: PMC1256015
- DOI: 10.1104/pp.105.068106
How to activate a plant gravireceptor. Early mechanisms of gravity sensing studied in characean rhizoids during parabolic flights
Abstract
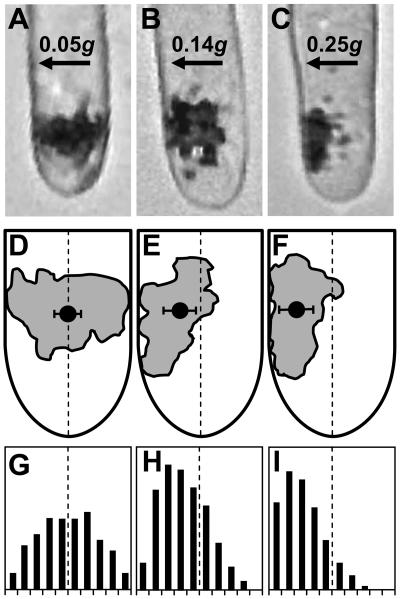
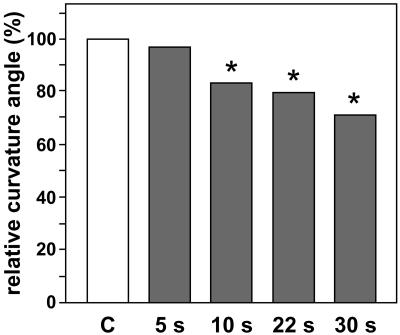
Early processes underlying plant gravity sensing were investigated in rhizoids of Chara globularis under microgravity conditions provided by parabolic flights of the A300-Zero-G aircraft and of sounding rockets. By applying centrifugal forces during the microgravity phases of sounding rocket flights, lateral accelerations of 0.14 g, but not of 0.05 g, resulted in a displacement of statoliths. Settling of statoliths onto the subapical plasma membrane initiated the gravitropic response. Since actin controls the positioning of statoliths and restricts sedimentation of statoliths in these cells, it can be calculated that lateral actomyosin forces in a range of 2 x 10(-14) n act on statoliths to keep them in place. These forces represent the threshold value that has to be exceeded by any lateral acceleration stimulus for statolith sedimentation and gravisensing to occur. When rhizoids were gravistimulated during parabolic plane flights, the curvature angles of the flight samples, whose sedimented statoliths became weightless for 22 s during the 31 microgravity phases, were not different from those of in-flight 1g controls. However, in ground control experiments, curvature responses were drastically reduced when the contact of statoliths with the plasma membrane was intermittently interrupted by inverting gravistimulated cells for less than 10 s. Increasing the weight of sedimented statoliths by lateral centrifugation did not enhance the gravitropic response. These results provide evidence that graviperception in characean rhizoids requires contact of statoliths with membrane-bound receptor molecules rather than pressure or tension exerted by the weight of statoliths.
Figures





Similar articles
-
Actomyosin-mediated statolith positioning in gravisensing plant cells studied in microgravity.J Plant Growth Regul. 2002 Jun;21(2):137-45. doi: 10.1007/s003440010052. Epub 2002 May 24. J Plant Growth Regul. 2002. PMID: 12016508
-
Hypergravity can reduce but not enhance the gravitropic response of Chara globularis protonemata.Protoplasma. 1998;204(3-4):145-54. doi: 10.1007/BF01280321. Protoplasma. 1998. PMID: 11542591
-
Statolith positioning by microfilaments in Chara rhizoids and protonemata.Adv Space Res. 1998;21(8-9):1183-9. doi: 10.1016/s0273-1177(97)00633-9. Adv Space Res. 1998. PMID: 11541370
-
Rhizoids and protonemata of characean algae: model cells for research on polarized growth and plant gravity sensing.Protoplasma. 2006 Dec;229(2-4):133-42. doi: 10.1007/s00709-006-0208-9. Epub 2006 Dec 16. Protoplasma. 2006. PMID: 17180494 Review.
-
Plant cells on earth and in space.Korean J Biol Sci. 2000 Sep;4(3):201-14. doi: 10.1080/12265071.2000.9647546. Korean J Biol Sci. 2000. PMID: 12760371 Review.
Cited by
-
Analysis of Graviresponse and Biological Effects of Vertical and Horizontal Clinorotation in Arabidopsis thaliana Root Tip.Plants (Basel). 2021 Apr 9;10(4):734. doi: 10.3390/plants10040734. Plants (Basel). 2021. PMID: 33918741 Free PMC article.
-
Direction of illumination controls gametophyte orientation in seedless plants and related algae.Plant Signal Behav. 2015;10(9):e1051277. doi: 10.1080/15592324.2015.1051277. Plant Signal Behav. 2015. PMID: 26237278 Free PMC article.
-
Gravity Signaling in Flowering Plant Roots.Plants (Basel). 2020 Sep 29;9(10):1290. doi: 10.3390/plants9101290. Plants (Basel). 2020. PMID: 33003550 Free PMC article. Review.
-
Surface tip-to-base Ca2+ and H+ ionic fluxes are involved in apical growth and graviperception of the Phycomyces stage I sporangiophore.Planta. 2012 Dec;236(6):1817-29. doi: 10.1007/s00425-012-1738-3. Epub 2012 Aug 22. Planta. 2012. PMID: 22910875
-
Electron tomographic characterization of a vacuolar reticulum and of six vesicle types that occupy different cytoplasmic domains in the apex of tip-growing Chara rhizoids.Planta. 2008 Apr;227(5):1101-14. doi: 10.1007/s00425-007-0684-y. Epub 2008 Jan 12. Planta. 2008. PMID: 18193275
References
-
- Björkman T (1988) Perception of gravity by plants. Adv Bot Res 15: 1–41 - PubMed
-
- Boonsirichai K, Guan C, Chen R, Masson PH (2002) Root gravitropism: an experimental tool to investigate basic cellular and molecular processes underlying mechanosensing and signal transmission in plants. Annu Rev Plant Physiol Plant Mol Biol 53: 421–447 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
