

CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis
- PMID: 16183837
- PMCID: PMC1255994
- DOI: 10.1104/pp.105.066928
CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis
Abstract
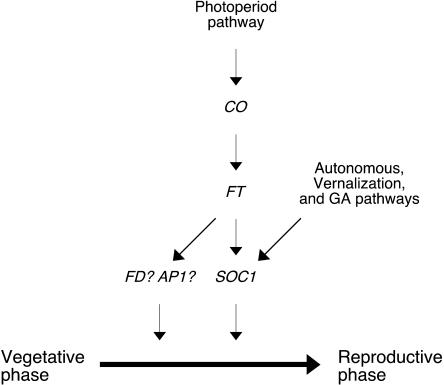
CONSTANS (CO) regulates flowering time by positively regulating expression of two floral integrators, FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1), in Arabidopsis (Arabidopsis thaliana). FT and SOC1 have been proposed to act in parallel pathways downstream of CO based on genetic analysis using weak ft alleles, since ft soc1 double mutants showed an additive effect in suppressing the early flowering of CO overexpressor plants. However, this genetic analysis was inconsistent with the sequential induction pattern of FT and SOC1 found in inducible CO overexpressor plants. Hence, to identify genetic interactions of CO, FT, and SOC1, we carried out genetic and expression analyses with a newly isolated T-DNA allele of FT, ft-10. We found that ft-10 almost completely suppressed the early flowering phenotype of CO overexpressor plants, whereas soc1-2 partially suppressed the phenotype, suggesting that FT is the major output of CO. Expression of SOC1 was altered in gain- or loss-of-function mutants of FT, whereas expression of FT remained unchanged in gain- or loss-of-function mutants of SOC1, suggesting that FT positively regulates SOC1 to promote flowering. In addition, inactivation of FT caused down-regulation of SOC1 even in plants overexpressing CO, indicating that FT is required for SOC1 induction by CO. Taken together, these data suggest that CO activates SOC1 through FT to promote flowering in Arabidopsis.
Figures





References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309: 1052–1056 - PubMed
-
- An H, Roussot C, Suarez-Lopez P, Corbesier L, Vincent C, Pineiro M, Hepworth S, Mouradov A, Justin S, Turnbull C, et al (2004) CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development 131: 3615–3626 - PubMed
-
- Banfield MJ, Brady RL (2000) The structure of Antirrhinum centroradialis protein (CEN) suggests a role as a kinase regulator. J Mol Biol 297: 1159–1170 - PubMed
-
- Bernier G (2004) Increased ethylene production in long days does not arise from activation of the ACS10 gene. Flowering Newsletter 38: 49–50
-
- Fabro G, Kovacs I, Pavet V, Szabados L, Alvarez ME (2004) Proline accumulation and AtP5CS2 gene activation are induced by plant-pathogen incompatible interactions in Arabidopsis. Mol Plant Microbe Interact 17: 343–350 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
