

Protein geranylgeranyltransferase I is involved in specific aspects of abscisic acid and auxin signaling in Arabidopsis
- PMID: 16183844
- PMCID: PMC1255991
- DOI: 10.1104/pp.105.065045
Protein geranylgeranyltransferase I is involved in specific aspects of abscisic acid and auxin signaling in Arabidopsis
Abstract
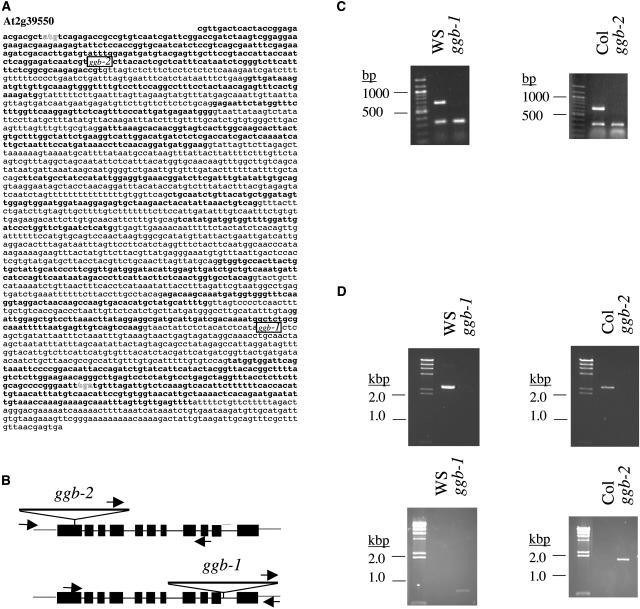
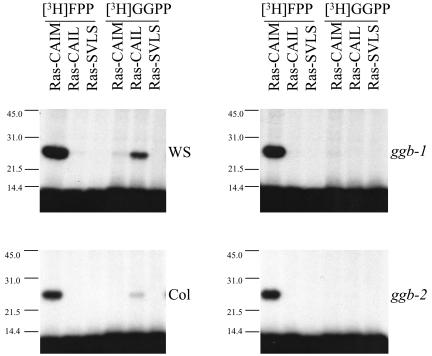
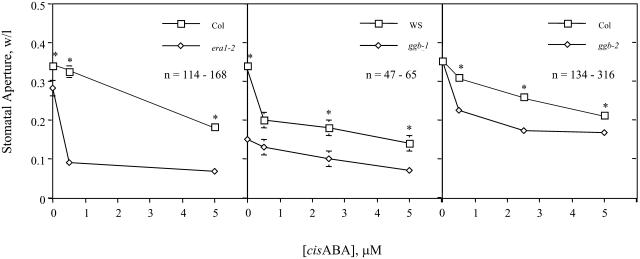
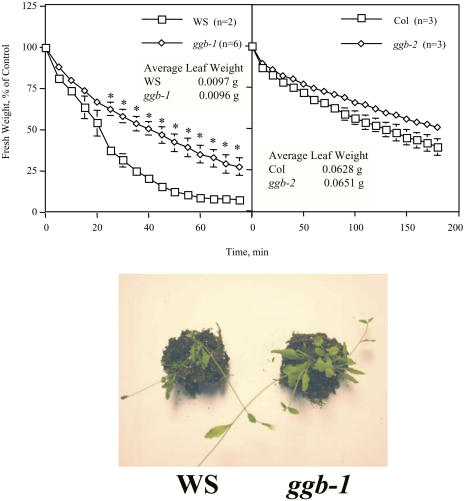
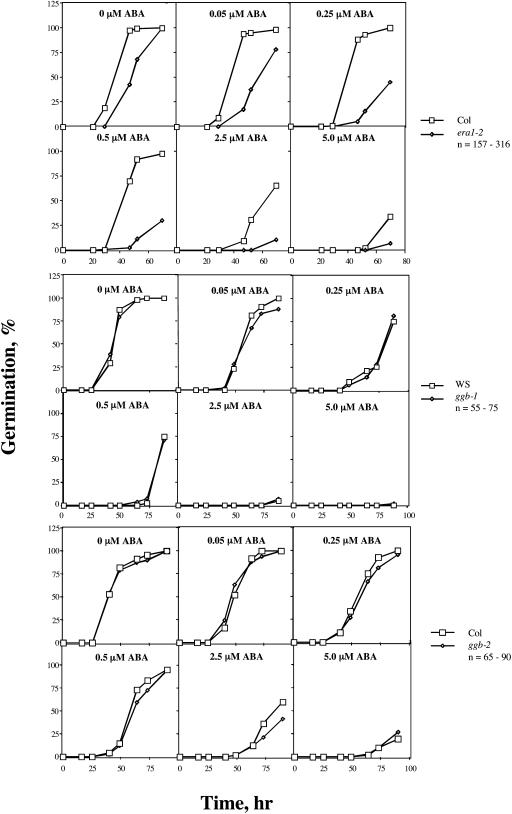
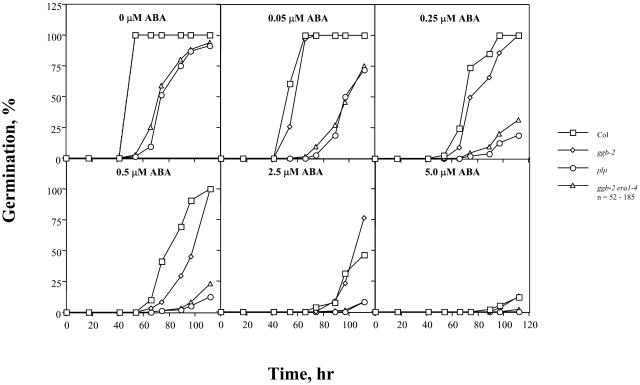
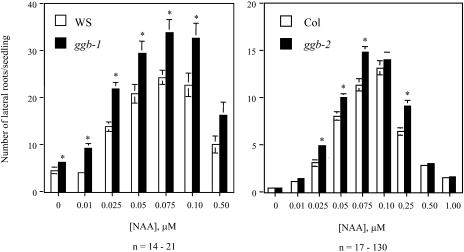
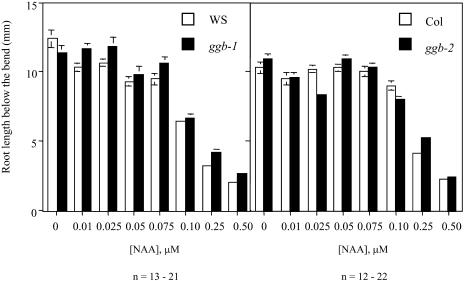
Arabidopsis (Arabidopsis thaliana) mutants lacking a functional ERA1 gene, which encodes the beta-subunit of protein farnesyltransferase (PFT), exhibit pleiotropic effects that establish roles for protein prenylation in abscisic acid (ABA) signaling and meristem development. Here, we report the effects of T-DNA insertion mutations in the Arabidopsis GGB gene, which encodes the beta-subunit of protein geranylgeranyltransferase type I (PGGT I). Stomatal apertures of ggb plants were smaller than those of wild-type plants at all concentrations of ABA tested, suggesting that PGGT I negatively regulates ABA signaling in guard cells. However, germination of ggb seeds in response to ABA was similar to the wild type. Lateral root formation in response to exogenous auxin was increased in ggb seedlings compared to the wild type, but no change in auxin inhibition of primary root growth was observed, suggesting that PGGT I is specifically involved in negative regulation of auxin-induced lateral root initiation. Unlike era1 mutants, ggb mutants exhibited no obvious developmental phenotypes. However, era1 ggb double mutants exhibited more severe developmental phenotypes than era1 mutants and were indistinguishable from plp mutants lacking the shared alpha-subunit of PFT and PGGT I. Furthermore, overexpression of GGB in transgenic era1 plants partially suppressed the era1 phenotype, suggesting that the relatively weak phenotype of era1 plants is due to partial redundancy between PFT and PGGT I. These results are discussed in the context of Arabidopsis proteins that are putative substrates of PGGT I.
Figures


Similar articles
-
The CaaX specificities of Arabidopsis protein prenyltransferases explain era1 and ggb phenotypes.BMC Plant Biol. 2010 Jun 18;10:118. doi: 10.1186/1471-2229-10-118. BMC Plant Biol. 2010. PMID: 20565889 Free PMC article.
-
The ABSCISIC ACID INSENSITIVE 3 (ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis.Plant J. 2003 Apr;34(1):67-75. doi: 10.1046/j.1365-313x.2003.01707.x. Plant J. 2003. PMID: 12662310
-
Identification, cloning and characterization of sis7 and sis10 sugar-insensitive mutants of Arabidopsis.BMC Plant Biol. 2008 Oct 14;8:104. doi: 10.1186/1471-2229-8-104. BMC Plant Biol. 2008. PMID: 18854047 Free PMC article.
-
Beyond the Usual Suspects: Physiological Roles of the Arabidopsis Amidase Signature (AS) Superfamily Members in Plant Growth Processes and Stress Responses.Biomolecules. 2021 Aug 13;11(8):1207. doi: 10.3390/biom11081207. Biomolecules. 2021. PMID: 34439873 Free PMC article. Review.
-
Protein farnesylation in plants: a greasy tale.Curr Opin Plant Biol. 1999 Oct;2(5):388-92. doi: 10.1016/s1369-5266(99)00010-2. Curr Opin Plant Biol. 1999. PMID: 10508754 Review.
Cited by
-
Protein Prenylation in Plant Stress Responses.Molecules. 2019 Oct 30;24(21):3906. doi: 10.3390/molecules24213906. Molecules. 2019. PMID: 31671559 Free PMC article. Review.
-
Identification of a novel abscisic acid-regulated farnesol dehydrogenase from Arabidopsis.Plant Physiol. 2010 Nov;154(3):1116-27. doi: 10.1104/pp.110.157784. Epub 2010 Aug 31. Plant Physiol. 2010. PMID: 20807998 Free PMC article.
-
Farnesylcysteine lyase is involved in negative regulation of abscisic acid signaling in Arabidopsis.Mol Plant. 2010 Jan;3(1):143-55. doi: 10.1093/mp/ssp091. Epub 2009 Nov 10. Mol Plant. 2010. PMID: 19969520 Free PMC article.
-
Differential effects of prenylation and s-acylation on type I and II ROPS membrane interaction and function.Plant Physiol. 2011 Feb;155(2):706-20. doi: 10.1104/pp.110.166850. Epub 2010 Dec 7. Plant Physiol. 2011. PMID: 21139084 Free PMC article.
-
Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes.Plant Physiol. 2011 Jul;156(3):1397-409. doi: 10.1104/pp.111.173427. Epub 2011 May 5. Plant Physiol. 2011. PMID: 21546455 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657; erratum Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Science 301: 1849 - PubMed
-
- Bergo MO, Leung GK, Ambroziak P, Otto JC, Casey PJ, Young SG (2000) Targeted inactivation of the isoprenylcysteine carboxyl methyltransferase gene causes mislocalization of K-Ras in mammalian cells. J Biol Chem 275: 17605–17610 - PubMed
-
- Bonetta D, Bayliss P, Sun S, Sage T, McCourt P (2000) Farnesylation is involved in meristem organization in Arabidopsis. Planta 211: 182–190 - PubMed
-
- Boyartchuk VL, Ashby MN, Rine J (1997) Modulation of Ras and a-factor function by carboxyl-terminal proteolysis. Science 275: 1796–1800 - PubMed
-
- Bracha K, Lavy M, Yalovsky S (2002) The Arabidopsis AtSTE24 is a CAAX protease with broad substrate specificity. J Biol Chem 277: 29856–29864 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
