

Quantitative modeling of Arabidopsis development
- PMID: 16183852
- PMCID: PMC1256009
- DOI: 10.1104/pp.105.060483
Quantitative modeling of Arabidopsis development
Abstract
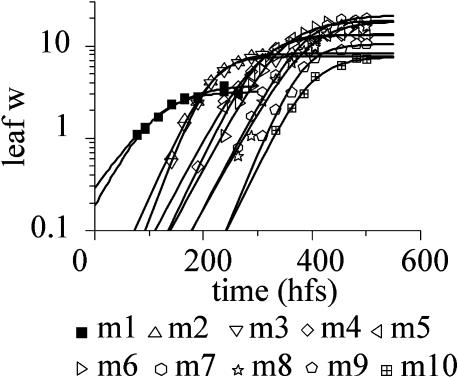
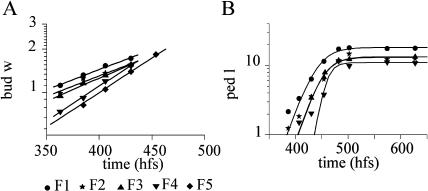
We present an empirical model of Arabidopsis (Arabidopsis thaliana), intended as a framework for quantitative understanding of plant development. The model simulates and realistically visualizes development of aerial parts of the plant from seedling to maturity. It integrates thousands of measurements, taken from several plants at frequent time intervals. These data are used to infer growth curves, allometric relations, and progression of shapes over time, which are incorporated into the final three-dimensional model. Through the process of model construction, we identify the key attributes required to characterize the development of Arabidopsis plant form over time. The model provides a basis for integrating experimental data and constructing mechanistic models.
Figures








References
-
- Bell AD (1986) Simulation of branching patterns in modular organisms. In JL Harper, BR Rosen, J White, eds, Growth and Form of Modular Organisms. Royal Society, London, pp 143–159
-
- Callos JD, Medford JI (1994) Organ positions and pattern formation in the shoot apex. Plant J 6: 1–7
-
- Chenu K, Franck N, Dauzat J, Lecoeur J (2004) Modelling the phenotypic variability of rosette architecture of Arabidopsis thaliana in several ecotypes and mutants in response to incident radiation. In C Godin, J Hanan, W Kurth, A Lacointe, A Takenaka, P Prusinkiewicz, T DeJong, C Beveridge, B Andrieu, eds, Proceedings of the 4th International Workshop on Functional-Structural Plant Models. Unité Mixte de Recherche AMAP, Montpellier, France, pp 360–364
-
- De Visser PHB, Marcelis LFM, van der Heijden GWAM, Angenent GC (2003) 3D digitization and modeling of flower mutants of Arabidopsis thaliana. In B-G Hu, M Jaeger, eds, Plant Growth Modeling and Applications. Tsinghua University Press and Springer, Beijing, pp 218–226
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
