

Dissection of Arp2/3 complex actin nucleation mechanism and distinct roles for its nucleation-promoting factors in Saccharomyces cerevisiae
- PMID: 16183906
- PMCID: PMC1456526
- DOI: 10.1534/genetics.105.040634
Dissection of Arp2/3 complex actin nucleation mechanism and distinct roles for its nucleation-promoting factors in Saccharomyces cerevisiae
Abstract
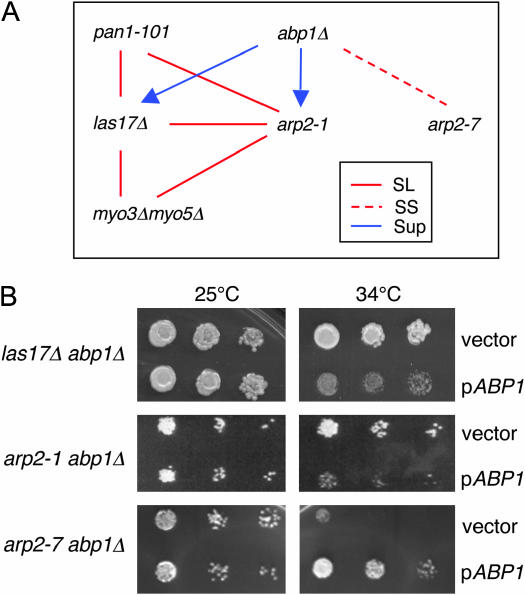
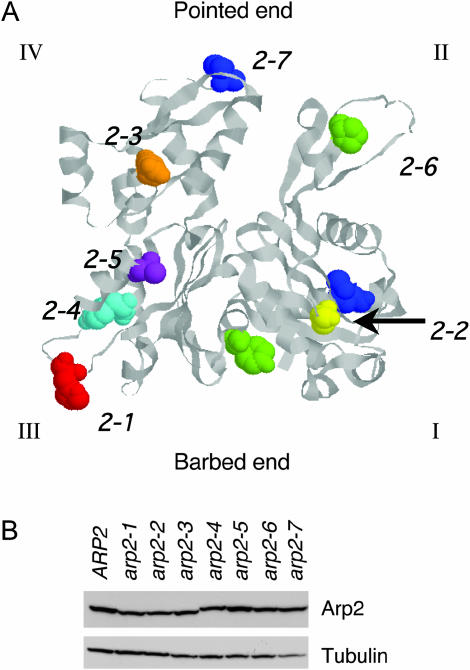
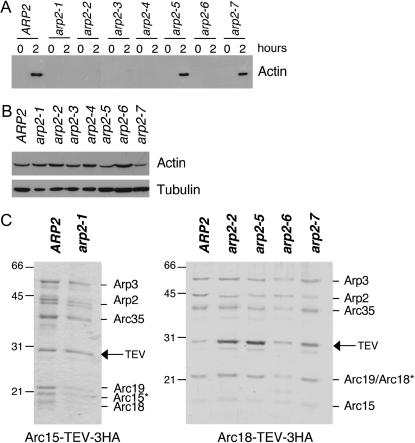
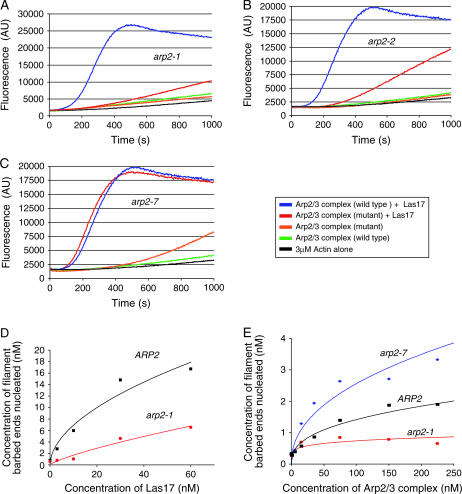
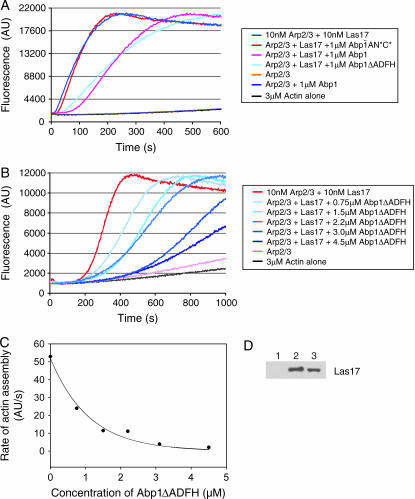
Actin nucleation by the Arp2/3 complex is under tight control, remaining inactive until stimulation by nucleation-promoting factors (NPFs). Although multiple NPFs are expressed in most cell types, little is known about how they are coordinated and whether they perform similar or distinct functions. We examined genetic relationships among the four S. cerevisiae NPFs. Combining las17delta with pan1-101 or myo3delta myo5delta was lethal at all temperatures, whereas combining pan1-101 with myo3delta myo5delta showed no genetic interaction and abp1delta partially suppressed las17delta. These data suggest that NPFs have distinct and overlapping functions in vivo. We also tested genetic interactions between each NPF mutant and seven different temperature-sensitive arp2 alleles and purified mutant Arp2/3 complexes to compare their activities. Two arp2 alleles with mutations at the barbed end were severely impaired in nucleation, providing the first experimental evidence that Arp2 nucleates actin at its barbed end in vitro and in vivo. Another arp2 allele caused partially unregulated ("leaky") nucleation in the absence of NPFs. Combining this mutant with a partially unregulated allele in a different subunit of Arp2/3 complex was lethal, suggesting that cells cannot tolerate high levels of unregulated activity. Genetic interactions between arp2 alleles and NPF mutants point to Abp1 having an antagonistic role with respect to other NPFs, possibly serving to attenuate their stronger activities. In support of this model, Abp1 binds strongly to Arp2/3 complex, yet has notably weak nucleation-promoting activity and inhibits Las17 activity on Arp2/3 complex in a dose-responsive manner.
Figures
References
-
- Beltzner, C. C., and T. D. Pollard, 2004. Identification of functionally important residues of Arp2/3 complex by analysis of homology models from diverse species. J. Mol. Biol. 336: 551–565. - PubMed
-
- Duncan, M. C., M. J. Cope, B. L. Goode, B. Wendland and D. G. Drubin, 2001. Yeast Eps15-like endocytic protein, Pan1p, activates the Arp2/3 complex. Nat. Cell Biol. 3: 687–690. - PubMed
-
- Goley, E. D., S. E. Rodenbusch, A. C. Martin and M. D. Welch, 2004. Critical conformational changes in the Arp2/3 complex are induced by nucleotide and nucleation promoting factor. Mol. Cell 16: 269–279. - PubMed
-
- Goode, B. L., 2002. Purification of yeast actin and actin-associated proteins. Methods Enzymol. 351: 433–441. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
