

The Rac activator Tiam1 controls tight junction biogenesis in keratinocytes through binding to and activation of the Par polarity complex
- PMID: 16186252
- PMCID: PMC2171536
- DOI: 10.1083/jcb.200502129
The Rac activator Tiam1 controls tight junction biogenesis in keratinocytes through binding to and activation of the Par polarity complex
Abstract
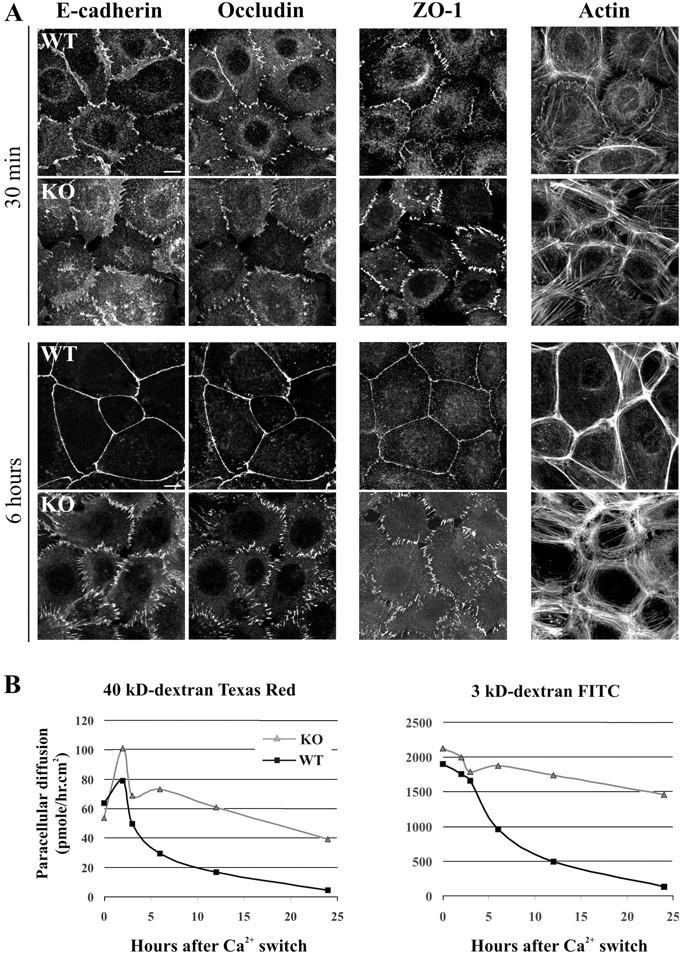
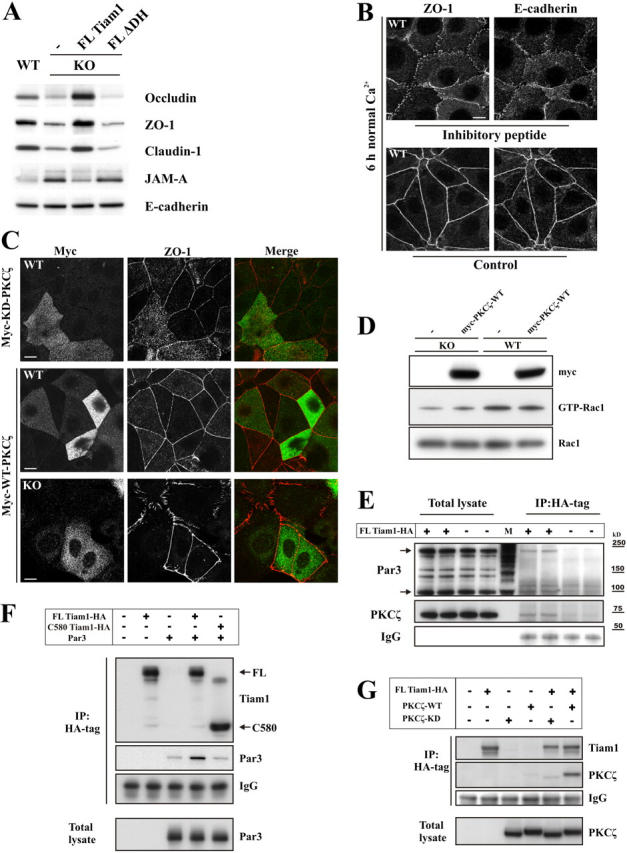
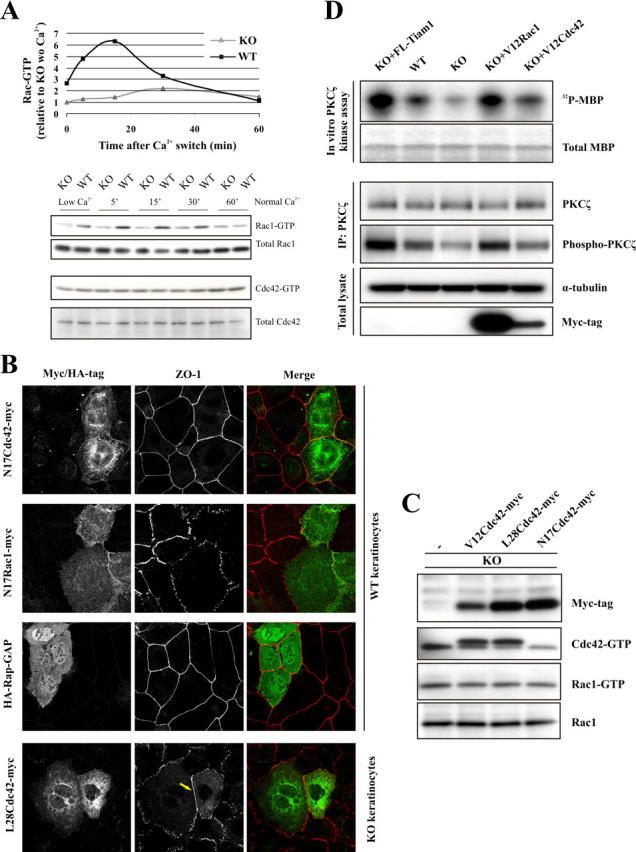
The GTPases Rac and Cdc42 play a pivotal role in the establishment of cell polarity by stimulating biogenesis of tight junctions (TJs). In this study, we show that the Rac-specific guanine nucleotide exchange factor Tiam1 (T-lymphoma invasion and metastasis) controls the cell polarity of epidermal keratinocytes. Similar to wild-type (WT) keratinocytes, Tiam1-deficient cells establish primordial E-cadherin-based adhesions, but subsequent junction maturation and membrane sealing are severely impaired. Tiam1 and V12Rac1 can rescue the TJ maturation defect in Tiam1-deficient cells, indicating that this defect is the result of impaired Tiam1-Rac signaling. Tiam1 interacts with Par3 and aPKCzeta, which are two components of the conserved Par3-Par6-aPKC polarity complex, and triggers biogenesis of the TJ through the activation of Rac and aPKCzeta, which is independent of Cdc42. Rac is activated upon the formation of primordial adhesions (PAs) in WT but not in Tiam1-deficient cells. Our data indicate that Tiam1-mediated activation of Rac in PAs controls TJ biogenesis and polarity in epithelial cells by association with and activation of the Par3-Par6-aPKC polarity complex.
Figures


References
-
- Chen, X., and I.G. Macara. 2005. Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat. Cell Biol. 7:262–269. - PubMed
-
- Fukuhara, A., K. Shimizu, T. Kawakatsu, T. Fukuhara, and Y. Takai. 2003. Involvement of nectin-activated Cdc42 small G protein in organization of adherens and tight junctions in Madin-Darby canine kidney cells. J. Biol. Chem. 278:51885–51893. - PubMed
-
- Gao, L., G. Joberty, and I.G. Macara. 2002. Assembly of epithelial tight junctions is negatively regulated by Par6. Curr. Biol. 12:221–225. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
