

Aurora A activates D-TACC-Msps complexes exclusively at centrosomes to stabilize centrosomal microtubules
- PMID: 16186253
- PMCID: PMC2171528
- DOI: 10.1083/jcb.200504097
Aurora A activates D-TACC-Msps complexes exclusively at centrosomes to stabilize centrosomal microtubules
Abstract
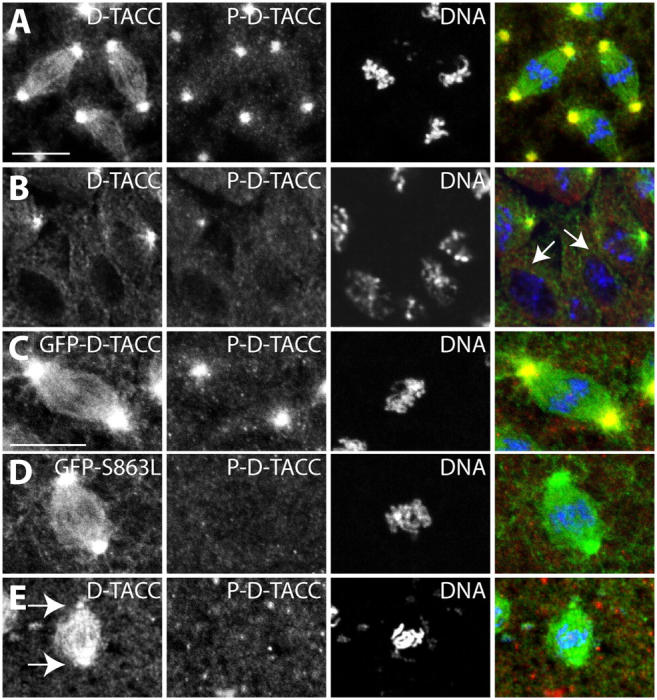
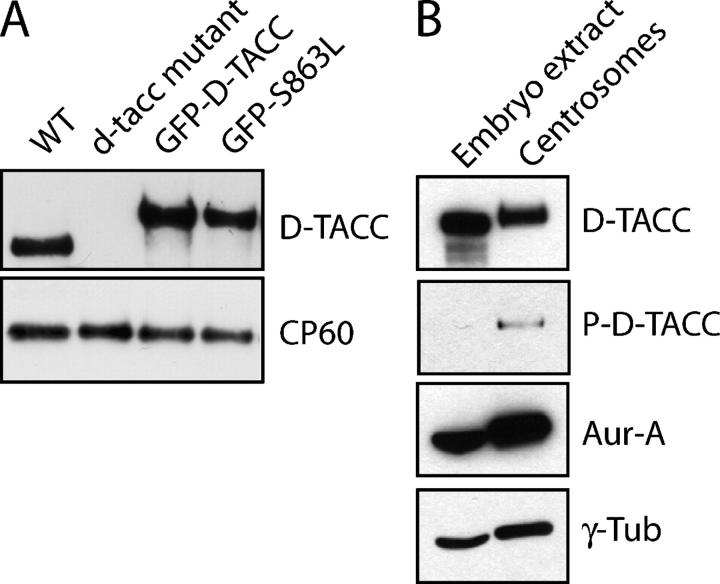
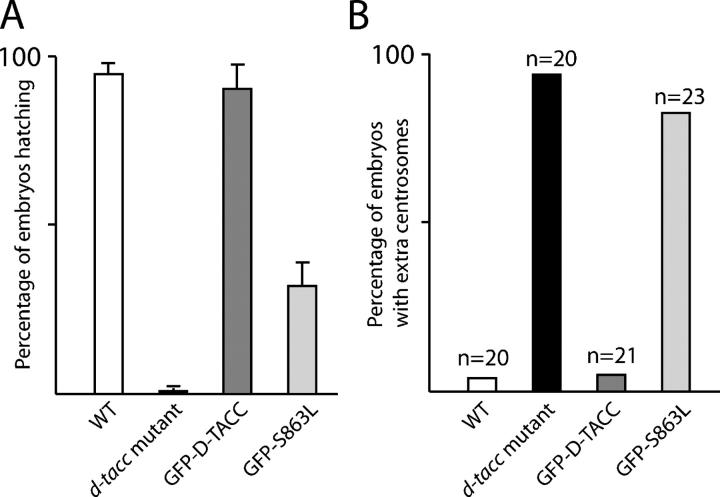
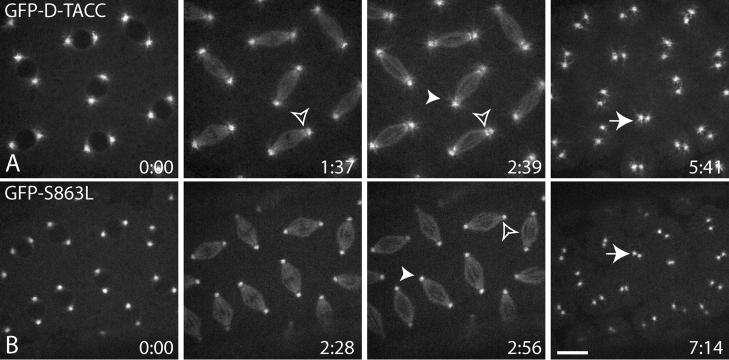
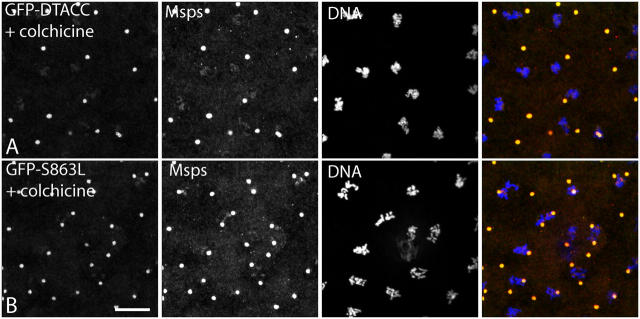
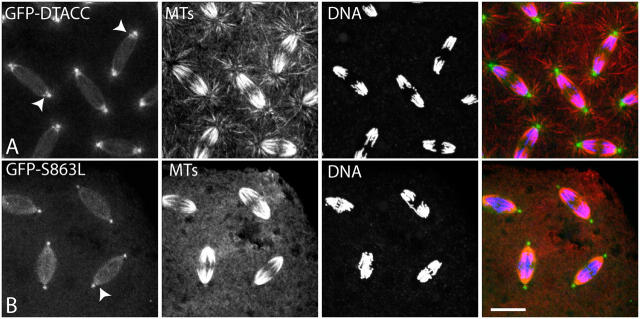
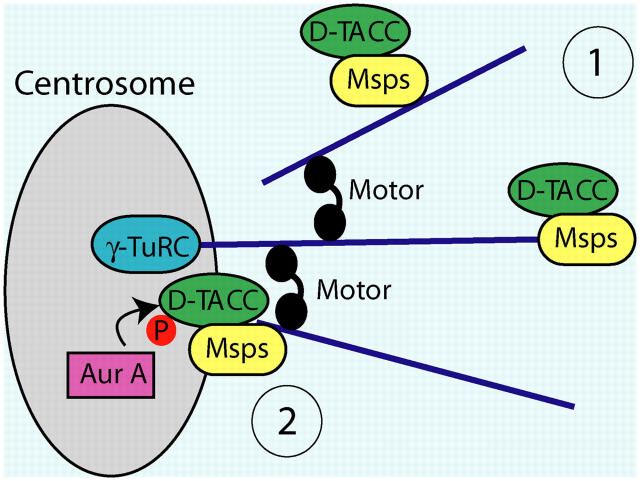
Centrosomes are the dominant sites of microtubule (MT) assembly during mitosis in animal cells, but it is unclear how this is achieved. Transforming acidic coiled coil (TACC) proteins stabilize MTs during mitosis by recruiting Minispindles (Msps)/XMAP215 proteins to centrosomes. TACC proteins can be phosphorylated in vitro by Aurora A kinases, but the significance of this remains unclear. We show that Drosophila melanogaster TACC (D-TACC) is phosphorylated on Ser863 exclusively at centrosomes during mitosis in an Aurora A-dependent manner. In embryos expressing only a mutant form of D-TACC that cannot be phosphorylated on Ser863 (GFP-S863L), spindle MTs are partially destabilized, whereas astral MTs are dramatically destabilized. GFP-S863L is concentrated at centrosomes and recruits Msps there but cannot associate with the minus ends of MTs. We propose that the centrosomal phosphorylation of D-TACC on Ser863 allows D-TACC-Msps complexes to stabilize the minus ends of centrosome-associated MTs. This may explain why centrosomes are such dominant sites of MT assembly during mitosis.
Figures
References
-
- Bellanger, J.M., and P. Gonczy. 2003. TAC-1 and ZYG-9 form a complex that promotes microtubule assembly in C. elegans embryos. Curr. Biol. 13:1488–1498. - PubMed
-
- Berdnik, D., and J.A. Knoblich. 2002. Drosophila Aurora-A is required for centrosome maturation and actin-dependent asymmetric protein localization during mitosis. Curr. Biol. 12:640–647. - PubMed
-
- Cassimeris, L. 1999. Accessory protein regulation of microtubule dynamics throughout the cell cycle. Curr. Opin. Cell Biol. 11:134–141. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
