

Adaptive sex differences in growth of pre-ovulation oocytes in a passerine bird
- PMID: 16188605
- PMCID: PMC1559945
- DOI: 10.1098/rspb.2005.3194
Adaptive sex differences in growth of pre-ovulation oocytes in a passerine bird
Abstract
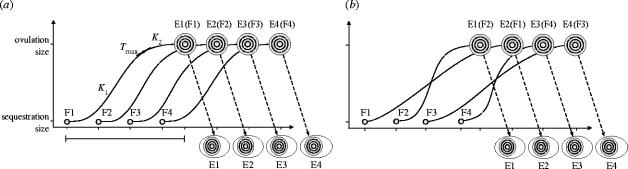
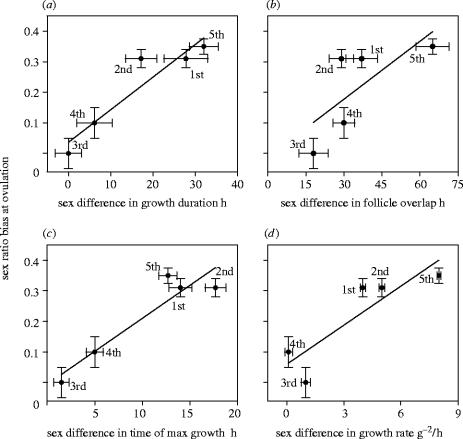
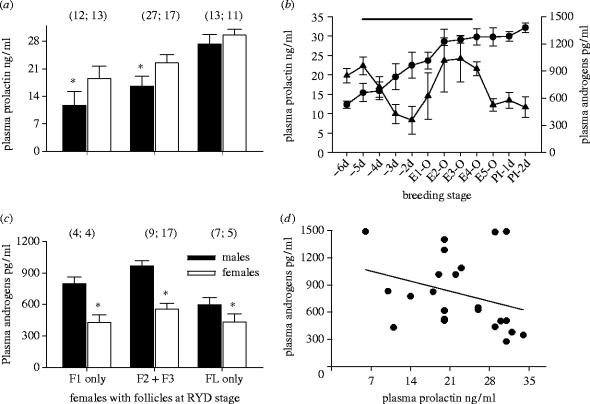
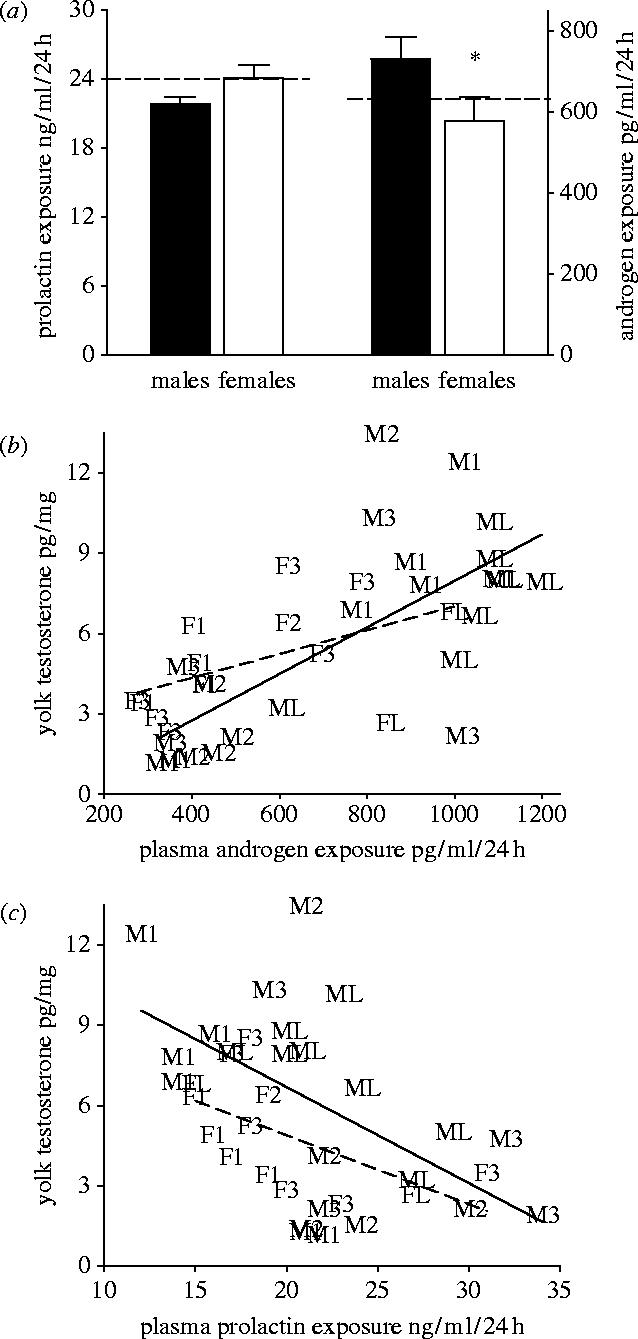
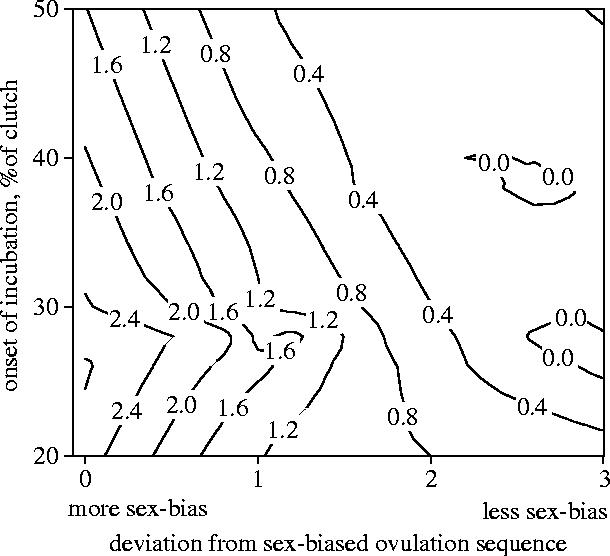
Maternal modification of offspring sex in birds has strong fitness consequences, however the mechanisms by which female birds can bias sex of their progeny in close concordance with the environment of breeding are not known. In recently established populations of house finches (Carpodacus mexicanus), breeding females lay a sex-biased sequence of eggs when ambient temperature causes early onset of incubation. We studied the mechanisms behind close association of incubation and sex-determination strategies in this species and discovered that pre-ovulation oocytes that produce males and females differed strongly in the temporal patterns of proliferation and growth. In turn, sex-specific exposure of oocytes to maternal secretion of prolactin and androgens produced distinct accumulation of maternal steroids in oocyte yolks in relation to oocyte proliferation order. These findings suggest that sex difference in oocyte growth and egg-laying sequence is an adaptive outcome of hormonal constraints imposed by the overlap of early incubation and oogenesis in this population, and that the close integration of maternal incubation, oocytes' sex-determination and growth might be under control of the same hormonal mechanism. We further document that population establishment and the evolution of these maternal strategies is facilitated by their strong effects on female and offspring fitness in a recently established part of the species range.
Figures
References
-
- Ankney C.D. Sex ratio varies with egg sequence in lesser snow geese. Auk. 1982;99:662–666.
-
- Astheimer L.B, Grau C.R. A comparison of yolk growth rates in seabird eggs. Ibis. 1990;132:380–394.
-
- Badyaev, A.V. in press Maternal inheritance and rapid evolution of sexual size dimorphism: passive effects or active strategies? Am. Nat.165 - PubMed
-
- Badyaev A.V, Martin T.E. Sexual dimorphism in relation to current selection in the house finch. Evolution. 2000;54:987–997. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources