

Endocytosis plays a critical role in proteolytic processing of the Hendra virus fusion protein
- PMID: 16188966
- PMCID: PMC1235849
- DOI: 10.1128/JVI.79.20.12643-12649.2005
Endocytosis plays a critical role in proteolytic processing of the Hendra virus fusion protein
Abstract
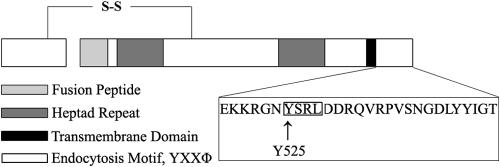
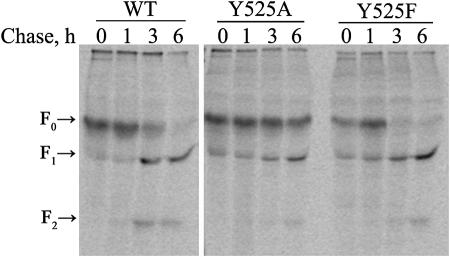
The Hendra virus fusion (F) protein is synthesized as a precursor protein, F(0), which is proteolytically processed to the mature form, F(1) + F(2). Unlike the case for the majority of paramyxovirus F proteins, the processing event is furin independent, does not require the addition of exogenous proteases, is not affected by reductions in intracellular Ca(2+), and is strongly affected by conditions that raise the intracellular pH (C. T. Pager, M. A. Wurth, and R. E. Dutch, J. Virol. 78:9154-9163, 2004). The Hendra virus F protein cytoplasmic tail contains a consensus motif for endocytosis, YXXPhi. To analyze the potential role of endocytosis in the processing and membrane fusion promotion of the Hendra virus F protein, mutation of tyrosine 525 to alanine (Hendra virus F Y525A) or phenylalanine (Hendra virus F Y525F) was performed. The rate of endocytosis of Hendra virus F Y525A was significantly reduced compared to that of the wild-type (wt) F protein, confirming the functional importance of the endocytosis motif. An intermediate level of endocytosis was observed for Hendra virus F Y525F. Surprisingly, dramatic reductions in the rate of proteolytic processing were observed for Hendra virus F Y525A, although initial transport to the cell surface was not affected. The levels of surface expression for both Hendra virus F Y525A and Hendra virus F Y525F were higher than that of the wt protein, and these mutants displayed enhanced syncytium formation. These results suggest that endocytosis is critically important for Hendra virus F protein cleavage, representing a new paradigm for proteolytic processing of paramyxovirus F proteins.
Figures
Similar articles
-
Cathepsin L is involved in proteolytic processing of the Hendra virus fusion protein.J Virol. 2005 Oct;79(20):12714-20. doi: 10.1128/JVI.79.20.12714-12720.2005. J Virol. 2005. PMID: 16188974 Free PMC article.
-
Subcellular localization and calcium and pH requirements for proteolytic processing of the Hendra virus fusion protein.J Virol. 2004 Sep;78(17):9154-63. doi: 10.1128/JVI.78.17.9154-9163.2004. J Virol. 2004. PMID: 15308711 Free PMC article.
-
Sequence motif upstream of the Hendra virus fusion protein cleavage site is not sufficient to promote efficient proteolytic processing.Virology. 2005 Oct 10;341(1):130-40. doi: 10.1016/j.virol.2005.07.004. Virology. 2005. PMID: 16083935
-
Role of N-linked glycosylation of the Hendra virus fusion protein.J Virol. 2005 Jun;79(12):7922-5. doi: 10.1128/JVI.79.12.7922-7925.2005. J Virol. 2005. PMID: 15919949 Free PMC article.
-
Residues in the hendra virus fusion protein transmembrane domain are critical for endocytic recycling.J Virol. 2012 Mar;86(6):3014-26. doi: 10.1128/JVI.05826-11. Epub 2012 Jan 11. J Virol. 2012. PMID: 22238299 Free PMC article.
Cited by
-
Henipavirus mediated membrane fusion, virus entry and targeted therapeutics.Viruses. 2012 Feb;4(2):280-308. doi: 10.3390/v4020280. Epub 2012 Feb 13. Viruses. 2012. PMID: 22470837 Free PMC article. Review.
-
Glycoprotein interactions in paramyxovirus fusion.Future Virol. 2009 Jul 1;4(4):335-351. doi: 10.2217/fvl.09.17. Future Virol. 2009. PMID: 20161127 Free PMC article.
-
Trafficking and Activation of Henipavirus, Parahenipavirus, and Henipa-like Virus Fusion Proteins.Viruses. 2025 Jun 19;17(6):866. doi: 10.3390/v17060866. Viruses. 2025. PMID: 40573457 Free PMC article. Review.
-
Third Helical Domain of the Nipah Virus Fusion Glycoprotein Modulates both Early and Late Steps in the Membrane Fusion Cascade.J Virol. 2020 Sep 15;94(19):e00644-20. doi: 10.1128/JVI.00644-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32669342 Free PMC article.
-
Analysis of cathepsin and furin proteolytic enzymes involved in viral fusion protein activation in cells of the bat reservoir host.PLoS One. 2015 Feb 23;10(2):e0115736. doi: 10.1371/journal.pone.0115736. eCollection 2015. PLoS One. 2015. PMID: 25706132 Free PMC article.
References
-
- Anderson, E. D., L. Thomas, J. S. Hayflick, and G. Thomas. 1993. Inhibition of HIV-1 gp160-dependent membrane fusion by a furin-directed alpha 1-antitrypsin variant. J. Biol. Chem. 268:24887-24891. - PubMed
-
- Begona Ruiz-Arguello, M., L. Gonzalez-Reyes, L. J. Calder, C. Palomo, D. Martin, M. J. Saiz, B. Garcia-Barreno, J. J. Skehel, and J. A. Melero. 2002. Effect of proteolytic processing at two distinct sites on shape and aggregation of an anchorless fusion protein of human respiratory syncytial virus and fate of the intervening segment. Virology 298:317-326. - PubMed
-
- Bolt, G., and I. R. Pedersen. 1998. The role of subtilisin-like proprotein convertases for cleavage of the measles virus fusion glycoprotein in different cell types. Virology 252:387-398. - PubMed
-
- Buchholz, U. J., S. Finke, and K. K. Conzelmann. 1999. Generation of bovine respiratory syncytial virus (BRSV) from cDNA: BRSV NS2 is not essential for virus replication in tissue culture, and the human RSV leader region acts as a functional BRSV genome promoter. J. Virol. 73:251-259. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
