

Low-dose adenovirus vaccine encoding chimeric hepatitis B virus surface antigen-human papillomavirus type 16 E7 proteins induces enhanced E7-specific antibody and cytotoxic T-cell responses
- PMID: 16188983
- PMCID: PMC1235816
- DOI: 10.1128/JVI.79.20.12807-12817.2005
Low-dose adenovirus vaccine encoding chimeric hepatitis B virus surface antigen-human papillomavirus type 16 E7 proteins induces enhanced E7-specific antibody and cytotoxic T-cell responses
Abstract
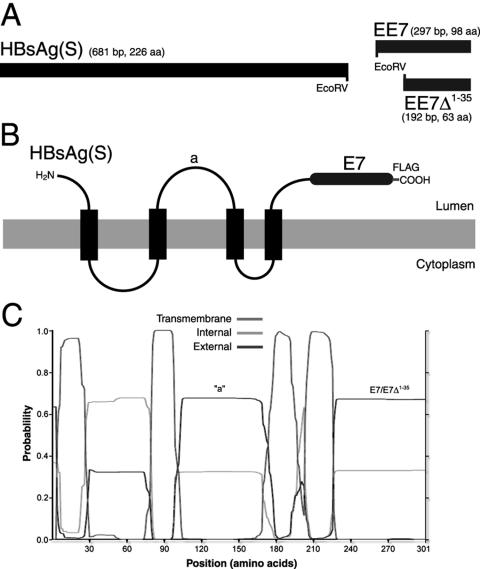
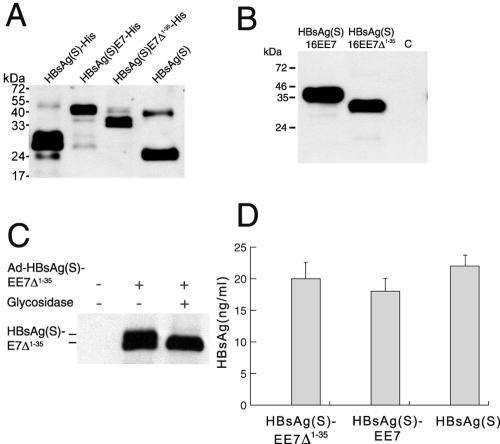
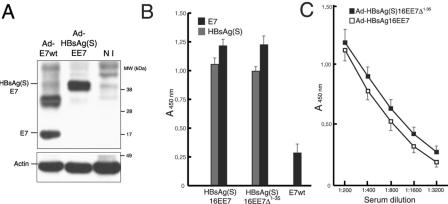
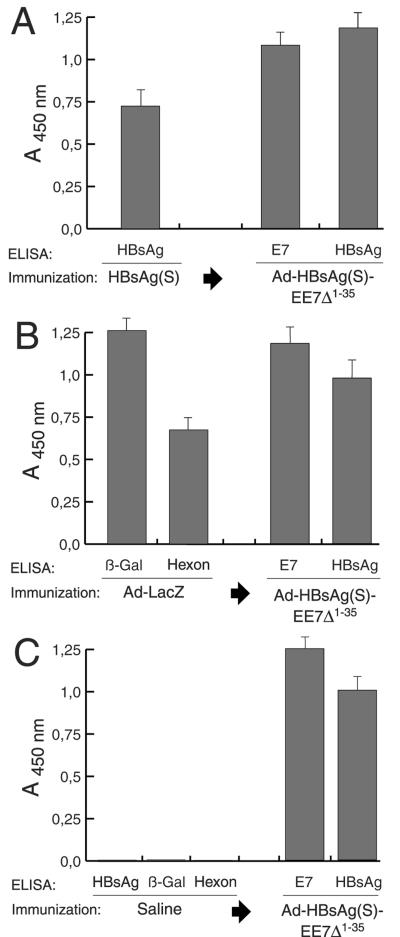
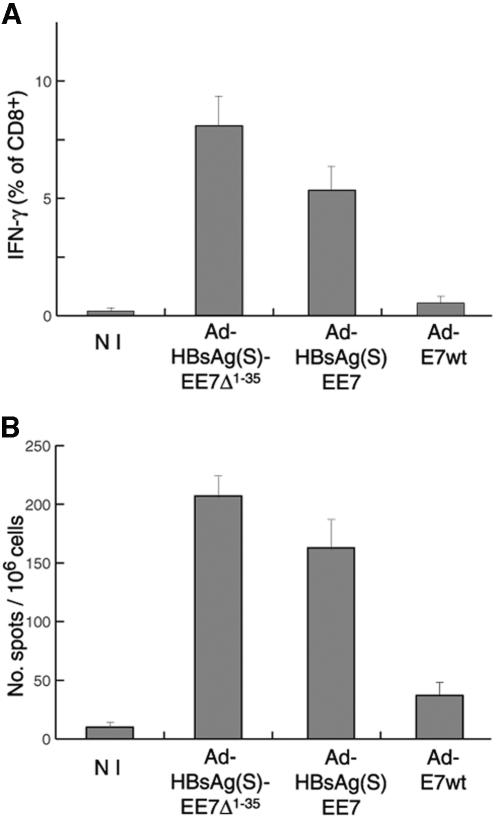
Induction of effective immune responses may help prevent cancer progression. Tumor-specific antigens, such as those of human papillomaviruses involved in cervical cancer, are targets with limited intrinsic immunogenicity. Here we show that immunization with low doses (10(6) infectious units/dose) of a recombinant human adenovirus type 5 encoding a fusion of the E7 oncoprotein of human papillomavirus type 16 to the carboxyl terminus of the surface antigen of hepatitis B virus (HBsAg) induces remarkable E7-specific humoral and cellular immune responses. The HBsAg/E7 fusion protein assembled efficiently into virus-like particles, which stimulated antibody responses against both carrier and foreign antigens, and evoked antigen-specific kill of an indicator cell population in vivo. Antibody and T-cell responses were significantly higher than those induced by a control adenovirus vector expressing wild-type E7. Such responses were not affected by preexisting immunity against either HBsAg or adenovirus. These data demonstrate that the presence of E7 on HBsAg particles does not interfere with particle secretion, as it occurs with bigger proteins fused to the C terminus of HBsAg, and results in enhancement of CD8(+)-mediated T-cell responses to E7. Thus, fusion to HBsAg is a convenient strategy for developing cervical cancer therapeutic vaccines, since it enhances the immunogenicity of E7 while turning it into an innocuous secreted fusion protein.
Figures


References
-
- Aichele, P., K. Brduscha-Riem, S. Oehen, B. Odermatt, R. M. Zinkernagel, H. Hengartner, and H. Pircher. 1997. Peptide antigen treatment of naive and virus-immune mice: antigen-specific tolerance versus immunopathology. Immunity 6:519-529. - PubMed
-
- Bontkes, H. J., T. D. de Gruijl, A. J. van den Muysenberg, R. H. Verheijen, M. J. Stukart, C. J. Meijer, R. J. Scheper, S. N. Stacey, M. F. Duggan Keen, P. L. Stern, S. Man, L. K. Borysiewicz, and J. M. Walboomers. 2000. Human papillomavirus type 16 E6/E7-specific cytotoxic T lymphocytes in women with cervical neoplasia. Int. J. Cancer. 88:92-98. - PubMed
-
- Davidson, E. J., R. L. Faulkner, P. Sehr, M. Pawlita, L. J. Smyth, D. J. Burt, A. E. Tomlinson, J. Hickling, H. C. Kitchener, and P. L. Stern. 2004. Effect of TA-CIN (HPV 16 L2E6E7) booster immunisation in vulval intraepithelial neoplasia patients previously vaccinated with TA-HPV (vaccinia virus encoding HPV 16/18 E6E7). Vaccine 22:2722-2729. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
