

Herpes simplex virus 1 envelopment follows two diverse pathways
- PMID: 16189007
- PMCID: PMC1235821
- DOI: 10.1128/JVI.79.20.13047-13059.2005
Herpes simplex virus 1 envelopment follows two diverse pathways
Abstract
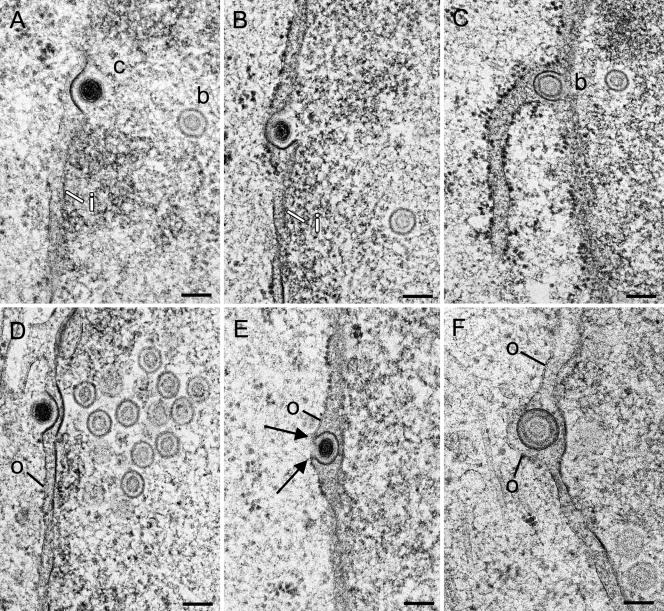
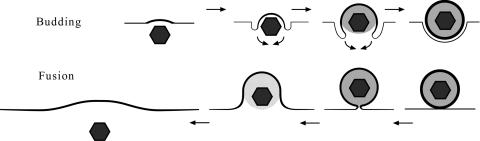
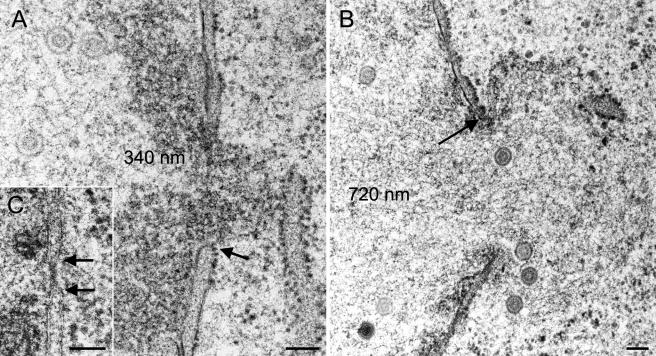
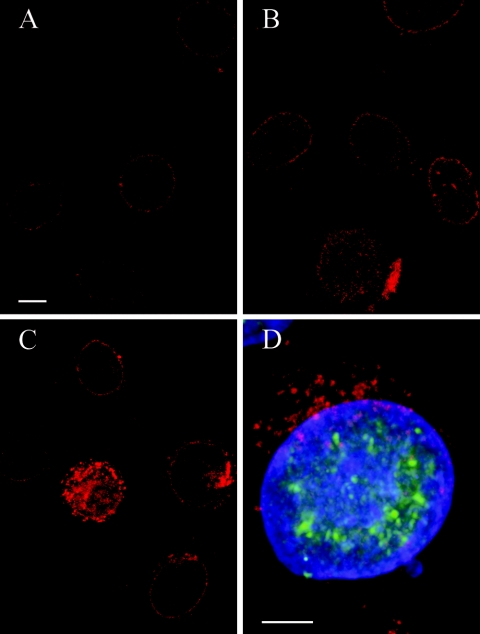
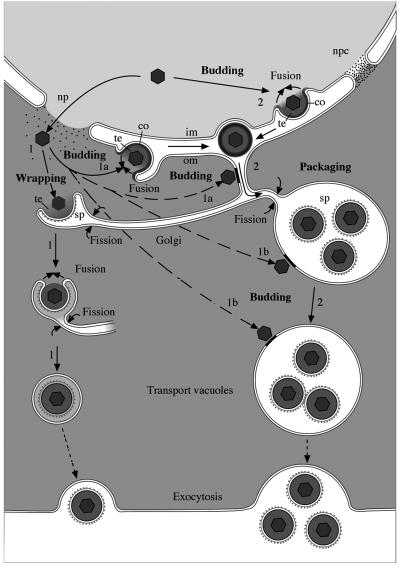
Herpesvirus envelopment is assumed to follow an uneconomical pathway including primary envelopment at the inner nuclear membrane, de-envelopment at the outer nuclear membrane, and reenvelopment at the trans-Golgi network. In contrast to the hypothesis of de-envelopment by fusion of the primary envelope with the outer nuclear membrane, virions were demonstrated to be transported from the perinuclear space to rough endoplasmic reticulum (RER) cisternae. Here we show by high-resolution microscopy that herpes simplex virus 1 envelopment follows two diverse pathways. First, nuclear envelopment includes budding of capsids at the inner nuclear membrane into the perinuclear space whereby tegument and a thick electron dense envelope are acquired. The substance responsible for the dense envelope is speculated to enable intraluminal transportation of virions via RER into Golgi cisternae. Within Golgi cisternae, virions are packaged into transport vacuoles containing one or several virions. Second, for cytoplasmic envelopment, capsids gain direct access from the nucleus to the cytoplasm via impaired nuclear pores. Cytoplasmic capsids could bud at the outer nuclear membrane, at membranes of RER, Golgi cisternae, and large vacuoles, and at banana-shaped membranous entities that were found to continue into Golgi membranes. Envelopes originating by budding at the outer nuclear membrane and RER membrane also acquire a dense substance. Budding at Golgi stacks, designated wrapping, results in single virions within small vacuoles that contain electron-dense substances between envelope and vacuolar membranes.
Figures
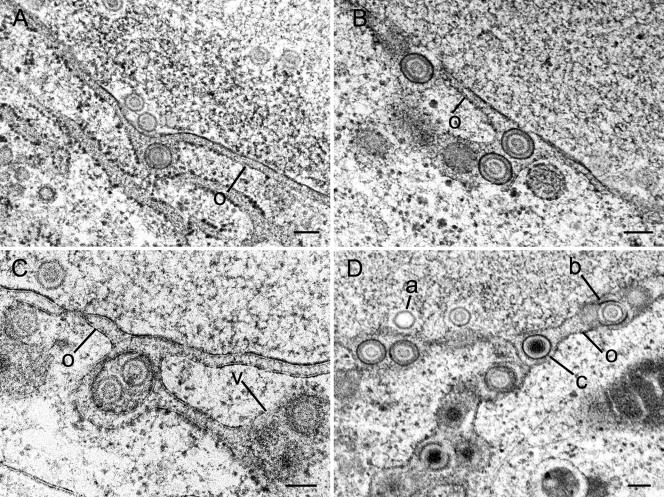
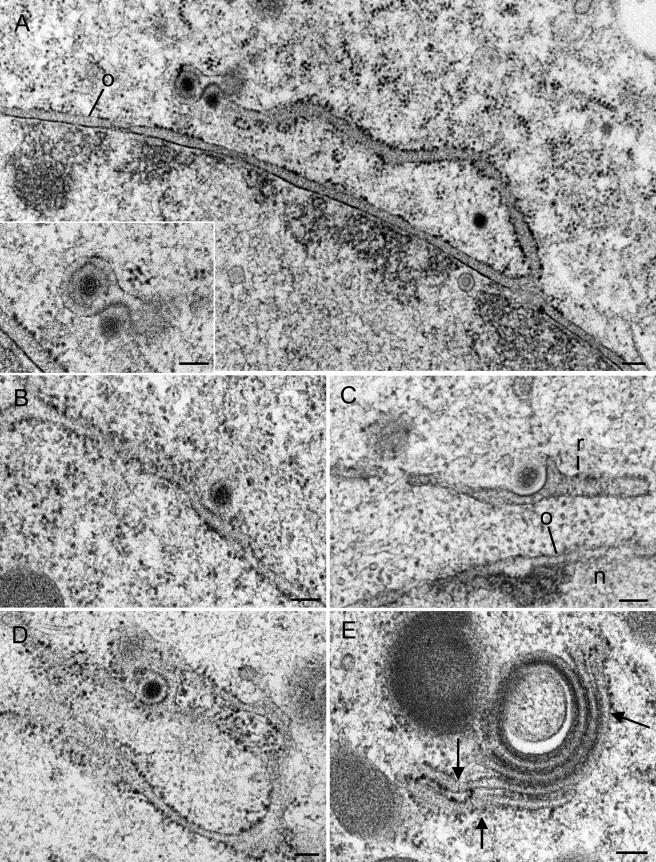
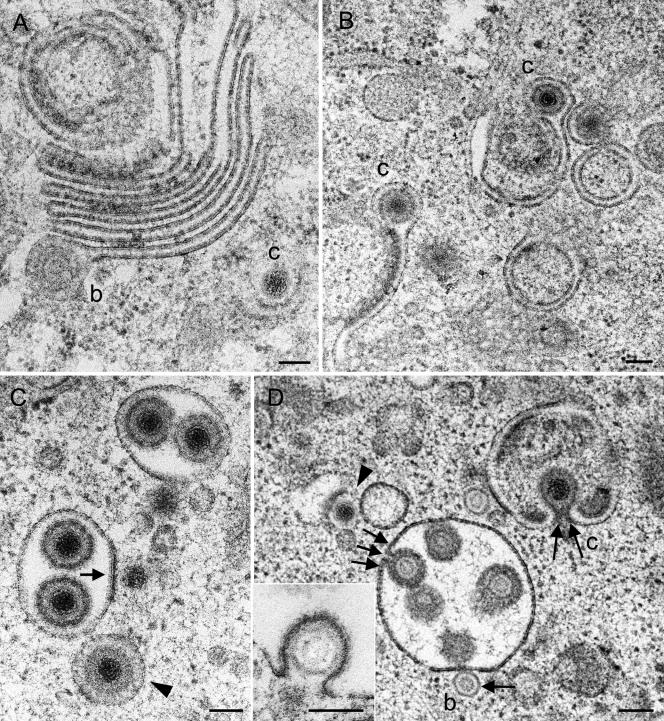
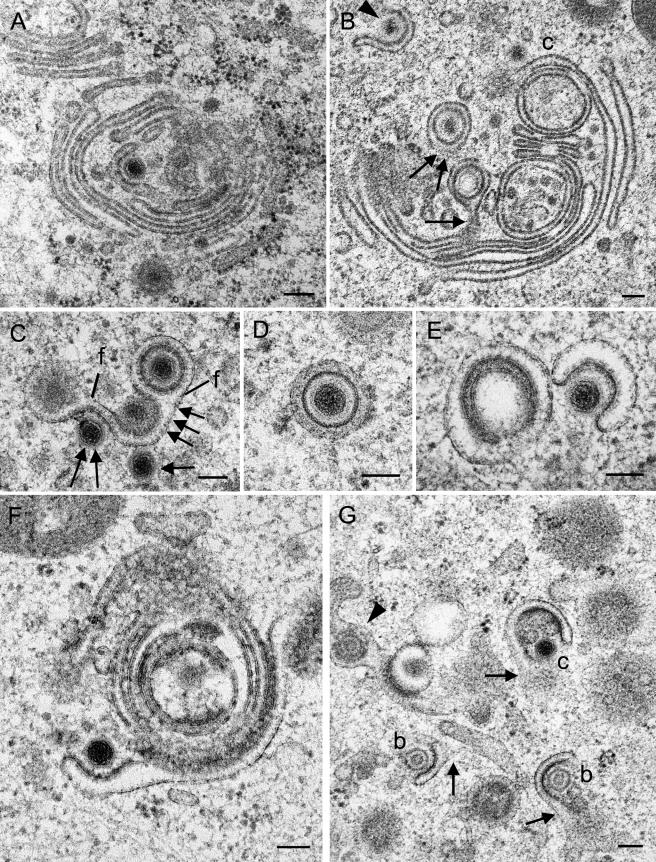
Comment in
-
Egress of alphaherpesviruses.J Virol. 2006 Feb;80(3):1610-1; author reply 1611-2. doi: 10.1128/JVI.80.3.1610-1612.2006. J Virol. 2006. PMID: 16415038 Free PMC article. No abstract available.
Similar articles
-
Impairment of nuclear pores in bovine herpesvirus 1-infected MDBK cells.J Virol. 2005 Jan;79(2):1071-83. doi: 10.1128/JVI.79.2.1071-1083.2005. J Virol. 2005. PMID: 15613336 Free PMC article.
-
Endoplasmic reticulum-to-Golgi transitions upon herpes virus infection.F1000Res. 2017 Oct 5;6:1804. doi: 10.12688/f1000research.12252.2. eCollection 2017. F1000Res. 2017. PMID: 30135710 Free PMC article.
-
Intra-nuclear localization of two envelope proteins, gB and gD, of herpes simplex virus.Arch Virol. 1996;141(3-4):505-24. doi: 10.1007/BF01718314. Arch Virol. 1996. PMID: 8645092
-
Herpesvirus Nuclear Egress across the Outer Nuclear Membrane.Viruses. 2021 Nov 24;13(12):2356. doi: 10.3390/v13122356. Viruses. 2021. PMID: 34960625 Free PMC article. Review.
-
Herpesvirus assembly: a tale of two membranes.Curr Opin Microbiol. 2006 Aug;9(4):423-9. doi: 10.1016/j.mib.2006.06.013. Epub 2006 Jun 30. Curr Opin Microbiol. 2006. PMID: 16814597 Review.
Cited by
-
Jumping over the fence: RNA nuclear export revisited.Nucleus. 2013 Mar-Apr;4(2):95-9. doi: 10.4161/nucl.24237. Epub 2013 Mar 1. Nucleus. 2013. PMID: 23528257 Free PMC article.
-
Crosstalk between nucleocytoplasmic trafficking and the innate immune response to viral infection.J Biol Chem. 2021 Jul;297(1):100856. doi: 10.1016/j.jbc.2021.100856. Epub 2021 Jun 29. J Biol Chem. 2021. PMID: 34097873 Free PMC article. Review.
-
Host and Viral Factors Involved in Nuclear Egress of Herpes Simplex Virus 1.Viruses. 2021 Apr 25;13(5):754. doi: 10.3390/v13050754. Viruses. 2021. PMID: 33923040 Free PMC article. Review.
-
Egress of alphaherpesviruses.J Virol. 2006 Feb;80(3):1610-1; author reply 1611-2. doi: 10.1128/JVI.80.3.1610-1612.2006. J Virol. 2006. PMID: 16415038 Free PMC article. No abstract available.
-
Herpes simplex virus type 1/adeno-associated virus hybrid vectors.Open Virol J. 2010 Jun 18;4:109-22. doi: 10.2174/1874357901004030109. Open Virol J. 2010. PMID: 20811580 Free PMC article.
References
-
- Avitabile, E., G. Lombardi, T. Gianni, M. Capri, and G. Campadelli-Fiume. 2004. Coexpression of UL20p and gK inhibits cell-cell fusion mediated by herpes simplex virus glycoproteins gD, gH-gL, and wild-type gB or an endocytosis-defective gB mutant and downmodulates their cell surface expression. J. Virol. 78:8015-8025. - PMC - PubMed
-
- Borchers, K., and M. Oezel. 1993. Simian agent 8 (SA8): morphogenesis and ultrastructure. Zentbl. Bakteriol. 279:526-536. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
