

doi: 10.1128/JVI.79.20.13173-13179.2005.
A role for herpesvirus entry mediator as the receptor for herpes simplex virus 1 entry into primary human trabecular meshwork cells
Affiliations
- PMID: 16189018
- PMCID: PMC1235852
- DOI: 10.1128/JVI.79.20.13173-13179.2005
Item in Clipboard
A role for herpesvirus entry mediator as the receptor for herpes simplex virus 1 entry into primary human trabecular meshwork cells
J Virol.
2005 Oct.
Abstract
The human eye is an important target for infection with herpes simplex virus 1 (HSV-1). Damage to cells forming the trabeculum of the eye by HSV-1 infection could contribute to the development of glaucoma, a major blinding disease. Primary cultures of human trabecular meshwork cells were used as an in vitro model to demonstrate the ability of HSV-1 to enter into and establish a productive infection of the trabeculum. Blocking of entry by anti-herpesvirus entry mediator (HVEM) antibody implicated HVEM as the major receptor for HSV-1 infection.
Figures
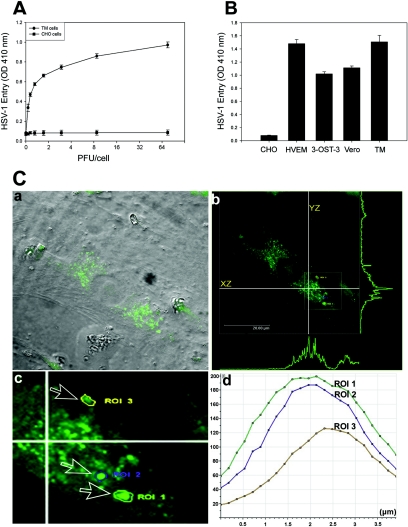
Analysis of HSV-1 entry in primary cultures of human TM cells. (A) Entry of human HSV-1 into cultured human TM cells. Cultured TM cells, along with wild-type CHO-K1 cells, were plated in 96-well plates and inoculated with twofold serial dilutions of β-galactosidase-expressing recombinant virus HSV-1 (KOS) gL86 at the PFU/cell indicated. After 6 h, the cells were washed, permeabilized, and incubated with o-nitrophenyl-β-d -galactopyranoside (ImmunoPure ONPG; Pierce) substrate for quantitation of β-galactosidase activity expressed from the input viral genome. The enzymatic activity was measured by spectrophotometer (Molecular Devices) at an optical density (OD) at 410 nm. Each value shown is the mean (± standard deviation) of three or more determinations. (B) Comparison of levels of HSV-1 entry into multiple cell types. Entry of HSV-1 into cultured human TM cells was compared to entry into naturally susceptible Vero cells and CHO-K1 cells expressing either HVEM or 3-OS HS as gD receptors. Approximately equal numbers of cells (20,000 cells per well, per Beckman counter estimate) were plated in 96-well plates at least 16 h prior to infection. All cell types were then treated with serial dilutions of β-galactosidase-expressing recombinant virus HSV-1 (KOS) gL86 for 6 h before the cells were washed and permeabilized, and substrate (ONPG) was added to measure the viral entry. The values shown (means ± standard deviations of triplicate determinations) represent the amount of reaction product detected spectrophotometrically at a single input dose of 40 PFU/cell. (C) Visualization of GFP-tagged HSV-1 (K26GFP) particles in cultured human TM cells by using confocal microscopy. Human TM cells were infected as described in the text. (a) Bright field image of cultured human TM cells overlaid with FITC fluorescent channel. Fluorescent viral capsids are seen in green. (b) Orthogonal section of the maximum projection of a z stack sliced at two different axes (xz and yz). Maximum fluorescence intensities are shown as green peaks. The inset shows the three ROI described in the text. (c) Enlarged view of the ROI (labeled and indicated by arrows). (d) Histogram of fluorescent intensities (y axis) obtained for z stacks (x axis). The total depth of the z stack was 4 μm.
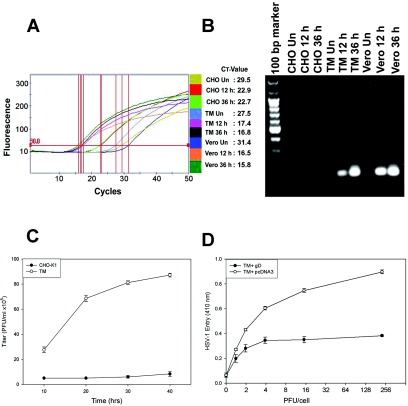
Entered HSV-1 replicates in human TM cells, and entry is sensitive to gD-mediated interference. (A) Real-time PCR analysis of entry. Fluorescence curve along with cycle threshold (CT) values are shown for each sample. The parameter CT is defined as the fractional cycle number at which the fluorescence passes the fixed threshold. Vero, TM, and CHO-K1 cells were infected with HSV-1. Total cell DNA was isolated 12 and 36 h postinfection and analyzed by RT-PCR. The reaction was carried out using the following conditions for the target HSV-1 (KOS) (gD). First, DNA was denatured at 95°C for 420 s, and the template was amplified for 50 cycles by denaturing DNA at 95°C for 30 s followed by annealing of primers at 58°C for 10 s and extension at 72°C for 16 s. After amplification, one cycle of melting curve from 60 to 95°C by a transition rate of 0.2°C/s, with continuous detection of fluorescence, was performed. Un, untreated. (B) Visualization of viral DNA. PCR products (20 μl) electrophoresed on 1.5% agarose (SeaKem LE agarose; Cambrex) Tris-acetate-EDTA (1×) gel containing ethidium bromide verified the amplification product (HSV-1 gD) of the predicted size (129 bp) with no primer dimer bands. Un, uninfected. (C) HSV-1 infection of TM cells leads to infectious yields of the virus. Confluent monolayers of TM and wild-type CHO-K1 cells were infected with HSV-1 at 0.01 PFU per cell for 90 min at 37°C. Inocula were harvested at the indicated times (10 to 40 h) postinfection. The infectious viral titers for each time point (PFU/ml) were determined in triplicate on Vero cells by plaque assay, which indicated that the viral titers in cultured corneal fibroblasts increased over the time period. Data represent the means ± standard deviations of results in triplicate wells from a representative experiment. The experiment was repeated three times with similar results. (D) Expression of HSV-1 gD in human TM cells interferes with HSV-1 entry. Cultured human TM cells were transfected with a plasmid expressing HSV-1 gD (pPEP99) (TM + gD) or with empty plasmid (pcDNA3) (TM + pcDNA3) as a control. Fourteen hours later, the cells were replated on 96-well plates and exposed to β-galactosidase-expressing HSV-1 (KOS) gL86. Six hours after inoculation, cells were lysed and β-galactosidase activities were determined as a measure of virus entry.
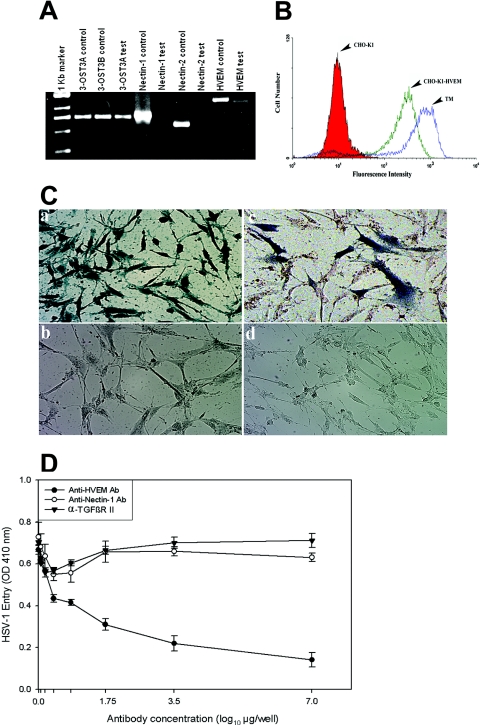
HVEM is the major mediator of entry into TM cells. (A) mRNAs specific to HVEM and 3-OST-3 but not to nectin-1 or nectin-2 were detected in TM cells. RT-PCR assays were performed. cDNAs were produced from total RNA isolated from the cells. PCR was performed using primers specific to each receptor (indicated at top). Either the expression plasmids for each receptor (indicated as controls) or cDNA isolated from TM cells (indicated as tests) were used as templates. The products were separated by electrophoresis on an agarose gel and then stained with ethidium bromide. Amplification products specific to HVEM and 3-OST-3 but not to nectin-1 or nectin-2 were detected in cDNAs generated from TM cells. (B) Cell surface expression of HVEM in TM cells detected by fluorescence-activated cell sorter (FACS) analysis. Monolayers of cultured TM cells were incubated at 4°C for 30 min with anti-HVEM antibody (1:200 dilution). CHO-K1 cells stably expressing HVEM (CHO-K1-HVEM) and wild-type CHO-K1 cells were used as positive and negative controls, respectively. Cells were examined by FACS analysis after 30 min of incubation with secondary anti-rabbit immunoglobulin G antibody (1:500 dilution) conjugated with FITC. (C) TM cells are resistant to BHV-1 entry. Cultured human TM cells (panels a and b), TM cells transiently transfected with the nectin-1 expression plasmid pBG38 (panel c), or TM cells mock transfected with empty vector (panel d) were exposed to β-galactosidase-expressing recombinants (40 PFU/cell) of HSV-1 (panel a) and BHV-1 (panels b, c, and d). After 6 h of infection at 37°C, cells were washed three times with PBS, fixed and permeabilized, and incubated with X-Gal (Invitrogen), which yields an insoluble blue product upon hydrolysis by β-galactosidase. Microscopy was performed using a 20× objective of the inverted microscope (Axiovert 100 M; Zeiss). The slide book version 3.0 was used for images. Blue cells (representing viral entry) were seen as shown. (D) Anti-HVEM polyclonal antibody, but not anti-nectin-1 antibody, inhibits HSV-1 entry into cultured human TM. Cells plated in 96-well plates were incubated with twofold dilutions of the anti-HVEM antibody (Ab), anti-nectin-1 Ab, and a control with α-TGFβR II Ab for 1 h 30 min at room temperature. Cells were then challenged with equal doses of HSV-1 (KOS) gL86 prepared in PBS with 1% glucose and 0.1% heat-inactivated calf serum at 37°C. After 2 h 30 min, cells were washed one time with PBS and treated for a short period with citrate buffer. Finally, cells were washed three times with PBS and incubated for 4 h in PBS buffer at 37°C. The substrate, ImmunoPure ONPG (o-nitrophenyl-β-d -galactopyranoside), was prepared in PBS buffer with nonionic detergent (Igepal CA-630; Sigma), and β-galactosidase activity was read at an optical density (OD) at 410 nm. The concentrations of the antibody used are expressed in μg/well. The experiment was repeated three times with similar results.
References
-
- Amano, S., T. Oshika, Y. Kaji, J. Numaga, M. Matsubara, and M. Araie. 1999. Herpes simplex virus in the trabeculum of an eye with corneal endotheliitis. Am. J. Ophthalmol. 127:721-722. - PubMed
-
- Bustos, D. E., and S. S. Atherton. 2002. Detection of herpes simplex virus type-1 in human ciliary ganglia. Investig. Ophthalmol. Vis. Sci. 43:2244-2249. - PubMed
-
- Cocchi, F., L. Menotti, P. Mirandola, M. Lopez, and G. Campadelli-Fiume. 1998. The ectodomain of a novel member of the immunoglobulin subfamily related to the poliovirus receptor has the attributes of a bona fide receptor for herpes simplex virus types 1 and 2 in human cells. J. Virol. 72:9992-10002. - PMC - PubMed
-
- Eberle, F., P. Dubreuil, M. G. Mattei, E. Devilard, and M. Lopez. 1995. The human PRR2 gene, related to the human poliovirus receptor gene (PVR), is the true homolog of the murine MPH gene. Gene 159:267-272. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
