

Homo- and heterodimerization of APP family members promotes intercellular adhesion
- PMID: 16193067
- PMCID: PMC1276707
- DOI: 10.1038/sj.emboj.7600824
Homo- and heterodimerization of APP family members promotes intercellular adhesion
Erratum in
- EMBO J. 2006 Feb 8;25(3):653
Abstract
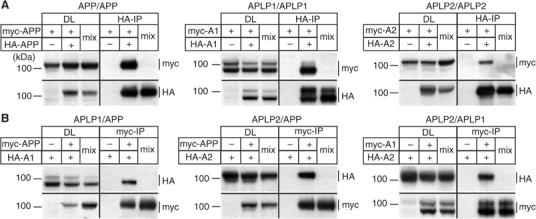
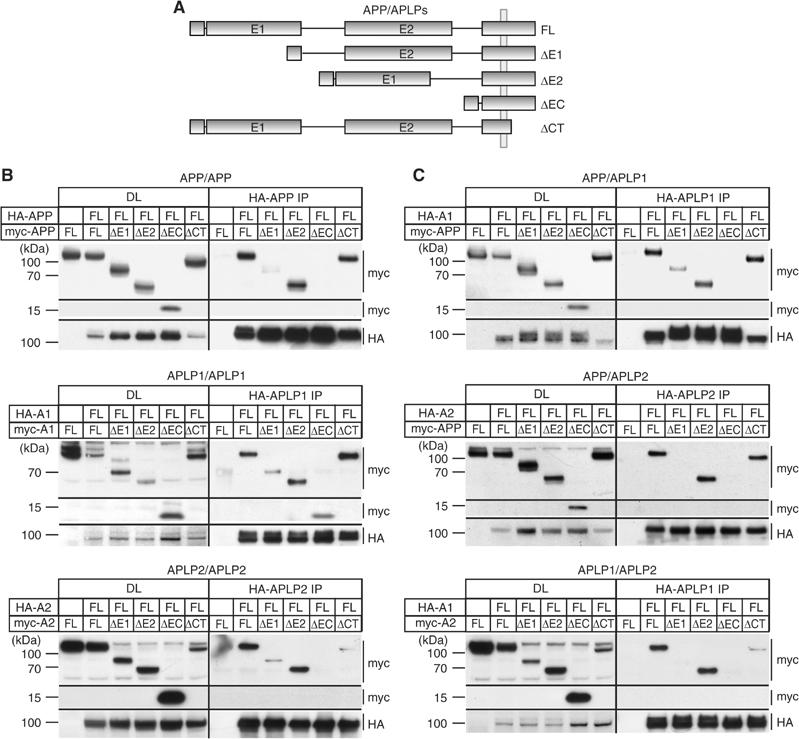
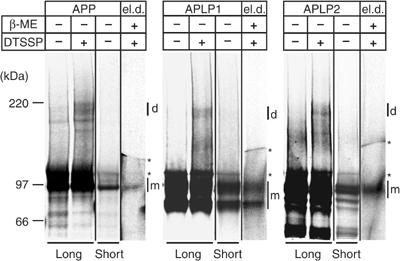
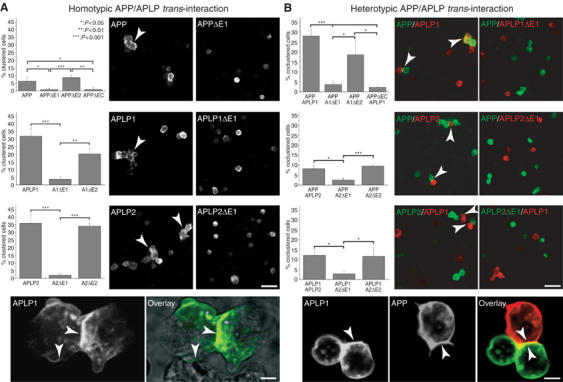
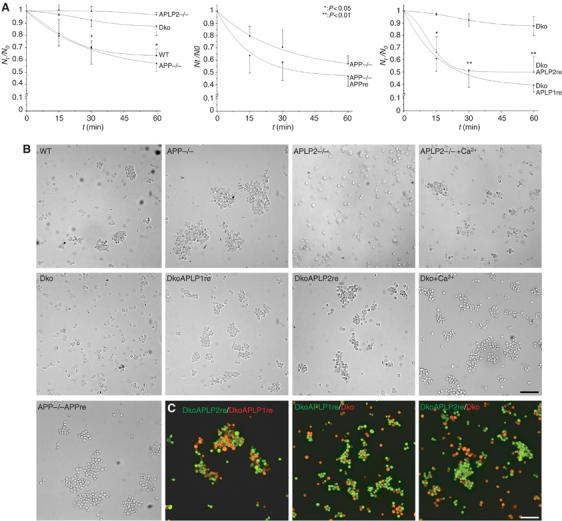
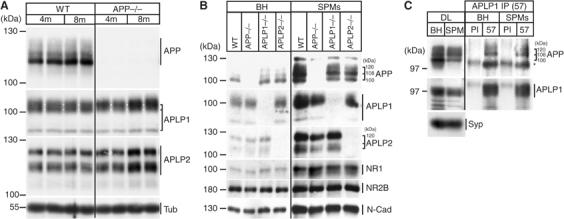
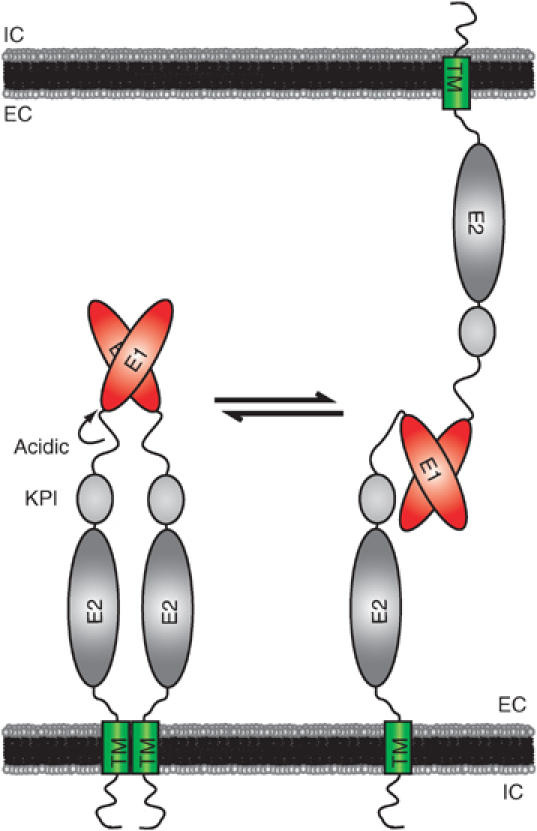
The amyloid precursor protein (APP) plays a central role in Alzheimer's disease, but its physiological function and that of its mammalian paralogs, the amyloid precursor-like proteins 1 and 2 (APLPs), is still poorly understood. APP has been proposed to form dimers, a process that could promote cell adhesion via trans-dimerization. We investigated the dimerization and cell adhesion properties of APP/APLPs and provide evidence that all three paralogs are capable of forming homo- and heterocomplexes. Moreover, we show that trans-interaction of APP family proteins promotes cell-cell adhesion in a homo- and heterotypic fashion and that endogenous APLP2 is required for cell-cell adhesion in mouse embryonic fibroblasts. We further demonstrate interaction of all the three APP family members in mouse brain, genetic interdependence, and molecular interaction of APP and APLPs in synaptically enriched membrane compartments. Together, our results provide evidence that homo- and heterocomplexes of APP/APLPs promote trans-cellular adhesion in vivo.
Figures

Similar articles
-
Subcellular localization and dimerization of APLP1 are strikingly different from APP and APLP2.J Cell Sci. 2009 Feb 1;122(Pt 3):368-77. doi: 10.1242/jcs.034058. Epub 2009 Jan 6. J Cell Sci. 2009. PMID: 19126676
-
Embryonic stem cell-derived neurons as a cellular system to study gene function: lack of amyloid precursor proteins APP and APLP2 leads to defective synaptic transmission.Stem Cells. 2008 Aug;26(8):2153-63. doi: 10.1634/stemcells.2008-0010. Epub 2008 Jun 5. Stem Cells. 2008. PMID: 18535156
-
The APP family of proteins: similarities and differences.Biochem Soc Trans. 2007 Apr;35(Pt 2):416-20. doi: 10.1042/BST0350416. Biochem Soc Trans. 2007. PMID: 17371289
-
The amyloid precursor protein and postnatal neurogenesis/neuroregeneration.Biochem Biophys Res Commun. 2006 Mar 3;341(1):1-5. doi: 10.1016/j.bbrc.2005.12.150. Epub 2006 Jan 5. Biochem Biophys Res Commun. 2006. PMID: 16406235 Review.
-
Functional interactions of APP with the apoE receptor family.J Neurochem. 2008 Sep;106(6):2263-71. doi: 10.1111/j.1471-4159.2008.05517.x. Epub 2008 Jun 28. J Neurochem. 2008. PMID: 18554321 Review.
Cited by
-
Advances in the pathogenesis of Alzheimer's disease: a re-evaluation of amyloid cascade hypothesis.Transl Neurodegener. 2012 Sep 21;1(1):18. doi: 10.1186/2047-9158-1-18. Transl Neurodegener. 2012. PMID: 23210692 Free PMC article.
-
Seizure susceptibility and mortality in mice that over-express amyloid precursor protein.Int J Clin Exp Pathol. 2008 Jan 1;1(2):157-68. Int J Clin Exp Pathol. 2008. PMID: 18784809 Free PMC article.
-
Amyloid Precursor Protein Dimerisation Reduces Neurite Outgrowth.Mol Neurobiol. 2019 Jan;56(1):13-28. doi: 10.1007/s12035-018-1070-4. Epub 2018 Apr 19. Mol Neurobiol. 2019. PMID: 29675574
-
Roles of amyloid precursor protein family members in neuroprotection, stress signaling and aging.Exp Brain Res. 2012 Apr;217(3-4):471-9. doi: 10.1007/s00221-011-2932-4. Epub 2011 Nov 16. Exp Brain Res. 2012. PMID: 22086493 Review.
-
Neuroprotective secreted amyloid precursor protein acts by disrupting amyloid precursor protein dimers.J Biol Chem. 2009 May 29;284(22):15016-25. doi: 10.1074/jbc.M808755200. Epub 2009 Mar 31. J Biol Chem. 2009. PMID: 19336403 Free PMC article.
References
-
- Aguzzi A, Haass C (2003) Games played by rogue proteins in prion disorders and Alzheimer's disease. Science 302: 814–818 - PubMed
-
- Annaert W, De Strooper B (2002) A cell biological perspective on Alzheimer's disease. Annu Rev Cell Dev Biol 18: 25–51 - PubMed
-
- Beher D, Elle C, Underwood J, Davis JB, Ward R, Karran E, Masters CL, Beyreuther K, Multhaup G (1999) Proteolytic fragments of Alzheimer's disease-associated presenilin 1 are present in synaptic organelles and growth cone membranes of rat brain. J Neurochem 72: 1564–1573 - PubMed
-
- Beher D, Hesse L, Masters CL, Multhaup G (1996) Regulation of amyloid protein precursor (APP) binding to collagen and mapping of the binding sites on APP and collagen type I. J Biol Chem 271: 1613–1620 - PubMed
-
- Coulson EJ, Paliga K, Beyreuther K, Masters CL (2000) What the evolution of the amyloid protein precursor supergene family tells us about its function. Neurochem Int 36: 175–184 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
