

Identification of GutQ from Escherichia coli as a D-arabinose 5-phosphate isomerase
- PMID: 16199563
- PMCID: PMC1251629
- DOI: 10.1128/JB.187.20.6936-6942.2005
Identification of GutQ from Escherichia coli as a D-arabinose 5-phosphate isomerase
Abstract
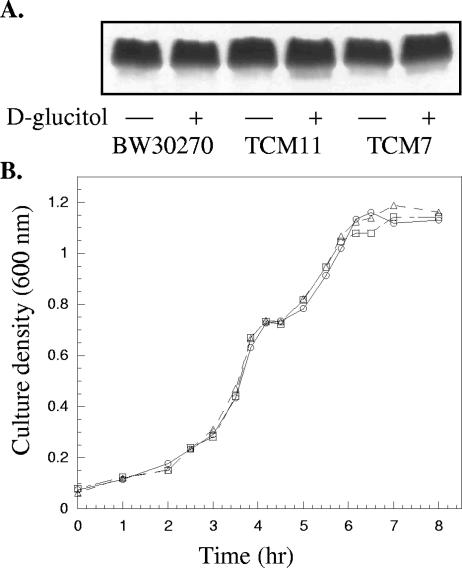
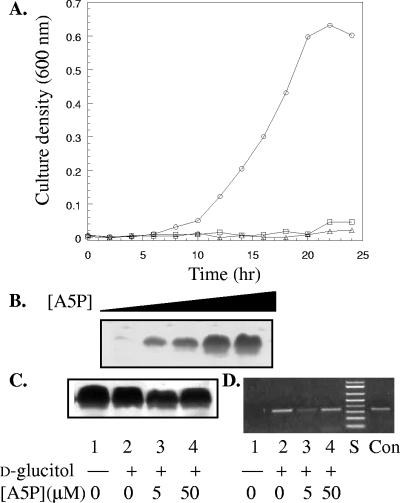
The glucitol operon (gutAEBDMRQ) of Escherichia coli encodes a phosphoenolpyruvate:sugar phosphotransferase system that metabolizes the hexitol D-glucitol (sorbitol). The functions for all but the last gene, gutQ, have been previously assigned. The high sequence similarity between GutQ and KdsD, a D-arabinose 5-phosphate isomerase (API) from the 3-deoxy-D-manno-octulosonate (KDO)-lipopolysaccharide (LPS) biosynthetic pathway, suggested a putative activity, but its role within the context of the gut operon remained unclear. Accordingly, the enzyme was cloned, overexpressed, and characterized. Recombinant GutQ was shown to indeed be a second copy of API from the E. coli K-12 genome with biochemical properties similar to those of KdsD, catalyzing the reversible aldol-ketol isomerization between D-ribulose 5-phosphate (Ru5P) and D-arabinose 5-phosphate (A5P). Genomic disruptions of each API gene were constructed in E. coli K-12. TCM11[(deltakdsD)] was capable of sustaining essential LPS synthesis at wild-type levels, indicating that GutQ functions as an API inside the cell. The gut operon remained inducible in TCM7[(deltagutQ)], suggesting that GutQ is not directly involved in d-glucitol catabolism. The conditional mutant TCM15[(deltagutQdeltakdsD)] was dependent on exogenous A5P both for LPS synthesis/growth and for upregulation of the gut operon. The phenotype was suppressed by complementation in trans with a plasmid encoding a functional copy of GutQ or by increasing the amount of A5P in the medium. As there is no obvious obligatory role for GutQ in the metabolism of d-glucitol and there is no readily apparent link between D-glucitol metabolism and LPS biosynthesis, it is suggested that A5P is not only a building block for KDO biosynthesis but also may be a regulatory molecule involved in expression of the gut operon.
Figures

References
-
- Aldridge, P., M. Metzger, and K. Geider. 1997. Genetics of sorbitol metabolism in Erwinia amylovora and its influence on bacterial virulence. Mol. Gen. Genet. 256:611-619. - PubMed
-
- Bateman, A. 1997. The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem. Sci. 22:12-13. - PubMed
-
- Bigham, E. C., C. E. Gragg, W. R. Hall, J. E. Kelsey, W. R. Mallory, D. C. Richardson, C. Benedict, and P. H. Ray. 1984. Inhibition of arabinose 5-phosphate isomerase. An approach to the inhibition of bacterial lipopolysaccharide biosynthesis. J. Med. Chem. 27:717-726. - PubMed
-
- Birck, M. R., T. P. Holler, and R. W. Woodard. 2000. Identification of a slow tight-binding inhibitor of 3-deoxy-dmanno-octulosonic acid 8-phosphate synthase. J. Am. Chem. Soc. 122:9334-9335.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
