

Nucleotide excision repair or polymerase V-mediated lesion bypass can act to restore UV-arrested replication forks in Escherichia coli
- PMID: 16199565
- PMCID: PMC1251618
- DOI: 10.1128/JB.187.20.6953-6961.2005
Nucleotide excision repair or polymerase V-mediated lesion bypass can act to restore UV-arrested replication forks in Escherichia coli
Abstract
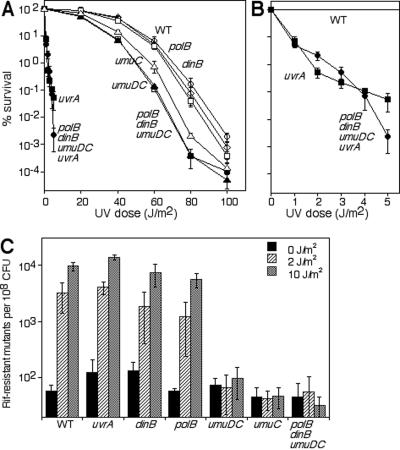
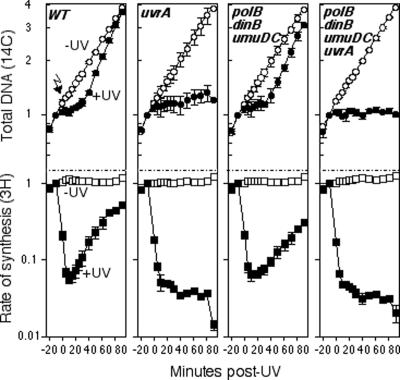
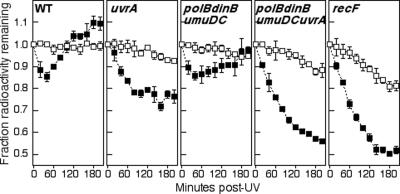
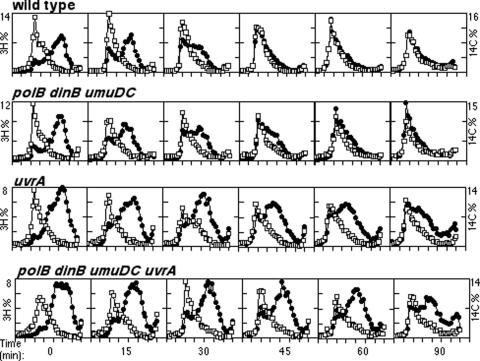
Nucleotide excision repair and translesion DNA synthesis are two processes that operate at arrested replication forks to reduce the frequency of recombination and promote cell survival following UV-induced DNA damage. While nucleotide excision repair is generally considered to be error free, translesion synthesis can result in mutations, making it important to identify the order and conditions that determine when each process is recruited to the arrested fork. We show here that at early times following UV irradiation, the recovery of DNA synthesis occurs through nucleotide excision repair of the lesion. In the absence of repair or when the repair capacity of the cell has been exceeded, translesion synthesis by polymerase V (Pol V) allows DNA synthesis to resume and is required to protect the arrested replication fork from degradation. Pol II and Pol IV do not contribute detectably to survival, mutagenesis, or restoration of DNA synthesis, suggesting that, in vivo, these polymerases are not functionally redundant with Pol V at UV-induced lesions. We discuss a model in which cells first use DNA repair to process replication-arresting UV lesions before resorting to mutagenic pathways such as translesion DNA synthesis to bypass these impediments to replication progression.
Figures
References
-
- Carty, M. P., C. W. Lawrence, and K. Dixon. 1996. Complete replication of plasmid DNA containing a single UV-induced lesion in human cell extracts. J. Biol. Chem. 271:9637-9647. - PubMed
-
- Chan, G. L., P. W. Doetsch, and W. A. Haseltine. 1985. Cyclobutane pyrimidine dimers and (6-4) photoproducts block polymerization by DNA polymerase I. Biochemistry 24:5723-5728. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
