

Regulating SWI/SNF subunit levels via protein-protein interactions and proteasomal degradation: BAF155 and BAF170 limit expression of BAF57
- PMID: 16199878
- PMCID: PMC1265786
- DOI: 10.1128/MCB.25.20.9016-9027.2005
Regulating SWI/SNF subunit levels via protein-protein interactions and proteasomal degradation: BAF155 and BAF170 limit expression of BAF57
Abstract
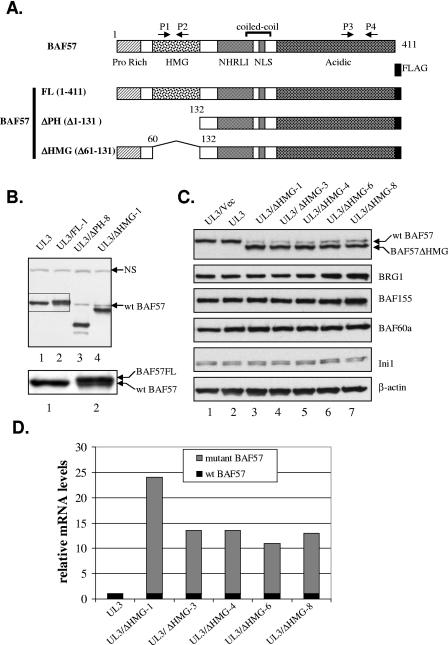
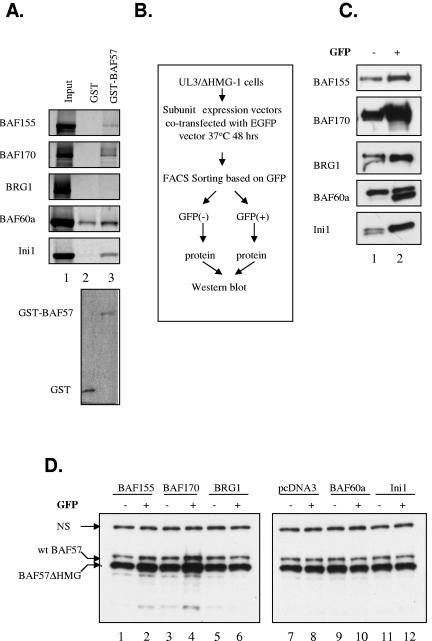
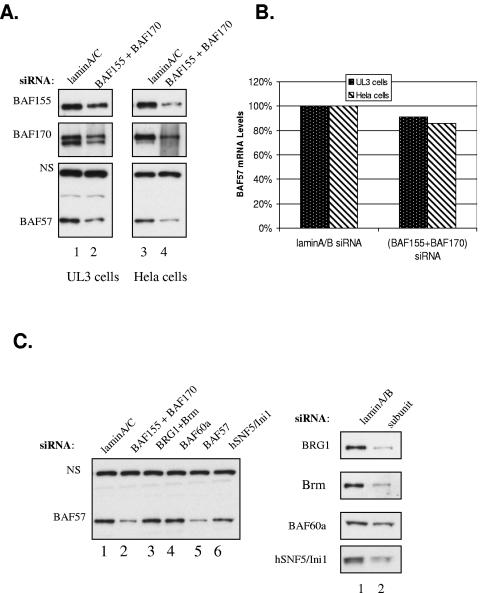
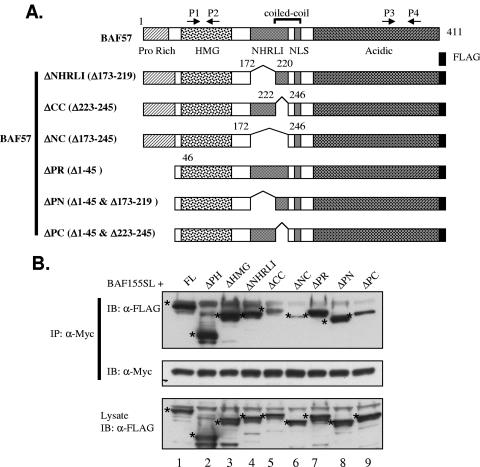
The mammalian SWI/SNF chromatin remodeling complex, whose function is of critical importance in transcriptional regulation, contains approximately 10 protein components. The expression levels of the core SWI/SNF subunits, including BRG1/Brm, BAF155, BAF170, BAF60, hSNF/Ini1, and BAF57, are stoichiometric, with few to no unbound molecules in the cell. Here we report that exogenous expression of the wild type or certain deletion mutants of BAF57, a key subunit that mediates the interaction between the remodeling complex and transcription factors, results in diminished expression of endogenous BAF57. This down-regulation process is mediated by an increase in proteasome-dependent degradation of the BAF57 protein. Furthermore, the protein levels of BAF155/170 dictate the maximum cellular amount of BAF57. We mapped the domains responsible for the interaction between BAF57 and BAF155 and demonstrated that protein-protein interactions between them play an important role in this regulatory process. These findings provide insights into the physiological mechanisms responsible for maintaining the proper stoichiometric levels of the protein components comprising multimeric enzyme complexes.
Figures


References
-
- Aasland, R., A. F. Stewart, and T. Gibson. 1996. The SANT domain: a putative DNA-binding domain in the SWI-SNF and ADA complexes, the transcriptional co-repressor N-CoR and TFIIIB. Trends Biochem. Sci. 21:87-88. - PubMed
-
- Aravind, L., and L. M. Iyer. 24July2002, posting date. The SWIRM domain: a conserved module found in chromosomal proteins points to novel chromatin-modifying activities. Genome Biol. 3:RESEARCH 0039. [Online.] http://genomebiology.com/2002/3/8/research0039. - PMC - PubMed
-
- Baker, K. M., G. Wei, A. E. Schaffner, and M. C. Ostrowski. 2003. Ets-2 and components of mammalian SWI/SNF form a repressor complex that negatively regulates the BRCA1 promoter. J. Biol. Chem. 278:17876-17884. - PubMed
-
- Boyer, L. A., R. R. Latek, and C. L. Peterson. 2004. The SANT domain: a unique histone-tail-binding module? Nat. Rev. Mol. Cell Biol. 5:158-163. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous