

Interaction between polyketide synthase and transporter suggests coupled synthesis and export of virulence lipid in M. tuberculosis
- PMID: 16201014
- PMCID: PMC1238737
- DOI: 10.1371/journal.ppat.0010002
Interaction between polyketide synthase and transporter suggests coupled synthesis and export of virulence lipid in M. tuberculosis
Abstract
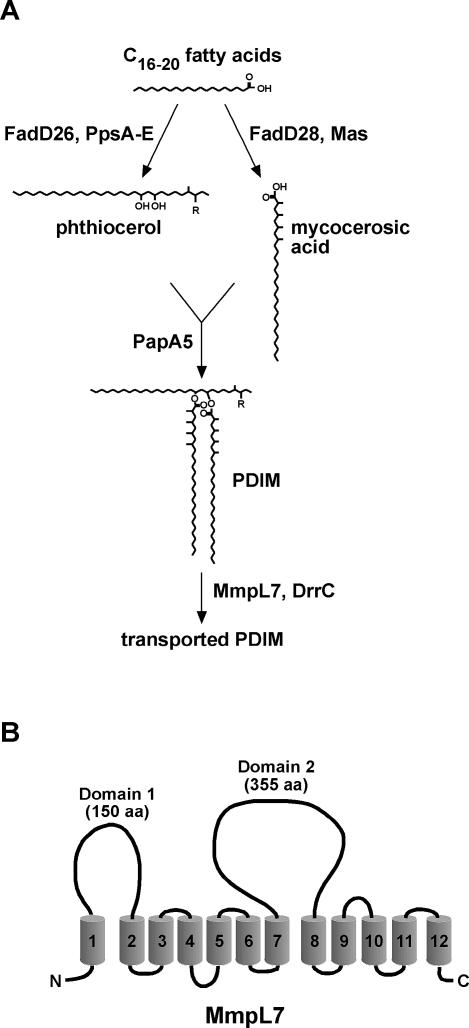
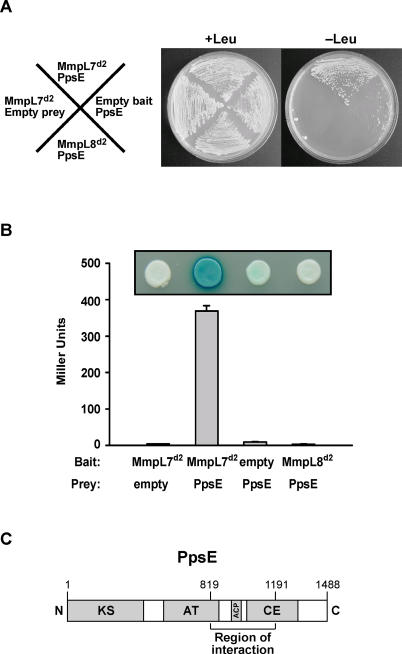
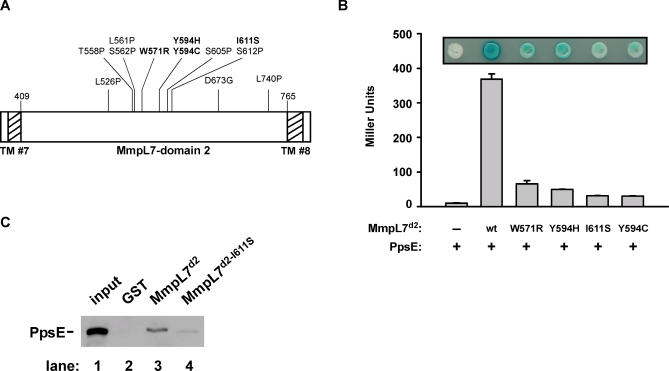
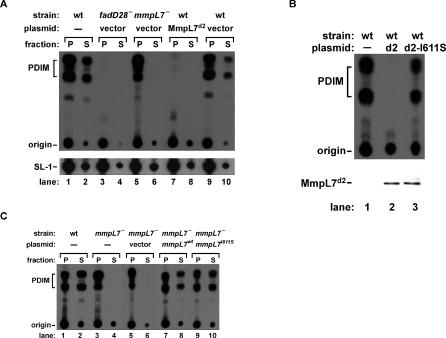
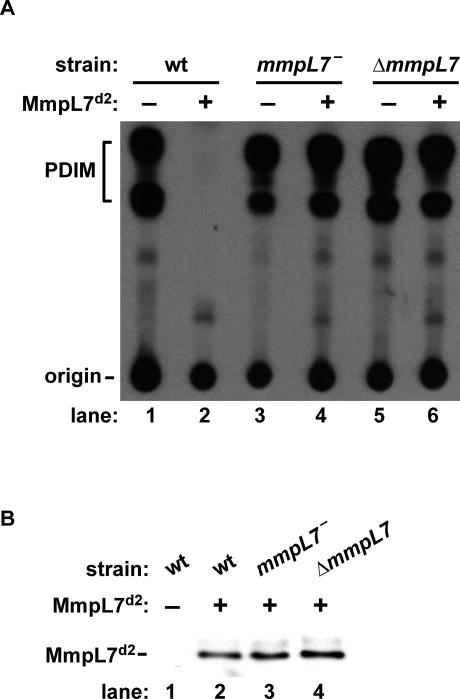
Virulent mycobacteria utilize surface-exposed polyketides to interact with host cells, but the mechanism by which these hydrophobic molecules are transported across the cell envelope to the surface of the bacteria is poorly understood. Phthiocerol dimycocerosate (PDIM), a surface-exposed polyketide lipid necessary for Mycobacterium tuberculosis virulence, is the product of several polyketide synthases including PpsE. Transport of PDIM requires MmpL7, a member of the MmpL family of RND permeases. Here we show that a domain of MmpL7 biochemically interacts with PpsE, the first report of an interaction between a biosynthetic enzyme and its cognate transporter. Overexpression of the interaction domain of MmpL7 acts as a dominant negative to PDIM synthesis by poisoning the interaction between synthase and transporter. This suggests that MmpL7 acts in complex with the synthesis machinery to efficiently transport PDIM across the cell membrane. Coordination of synthesis and transport may not only be a feature of MmpL-mediated transport in M. tuberculosis, but may also represent a general mechanism of polyketide export in many different microorganisms.
Conflict of interest statement
Figures

References
-
- McKinney JD, Jacobs WR, Jr, Bloom BR. Persisting problems in tuberculosis. In: Fauci A, Krause R, editors. Emerging infections. London: Academic Press; 1998. pp. 51–146.
-
- Brennan PJ, Nikaido H. The envelope of mycobacteria. Annu Rev Biochem. 1995;64:29–63. - PubMed
-
- Ng V, Zanazzi G, Timpl R, Talts JF, Salzer JL, et al. Role of the cell wall phenolic glycolipid-1 in the peripheral nerve predilection of Mycobacterium leprae . Cell. 2000;103:511–524. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
