

The analysis of interaural time differences in the chick brain stem
- PMID: 16202434
- PMCID: PMC1847356
- DOI: 10.1016/j.physbeh.2005.08.003
The analysis of interaural time differences in the chick brain stem
Abstract
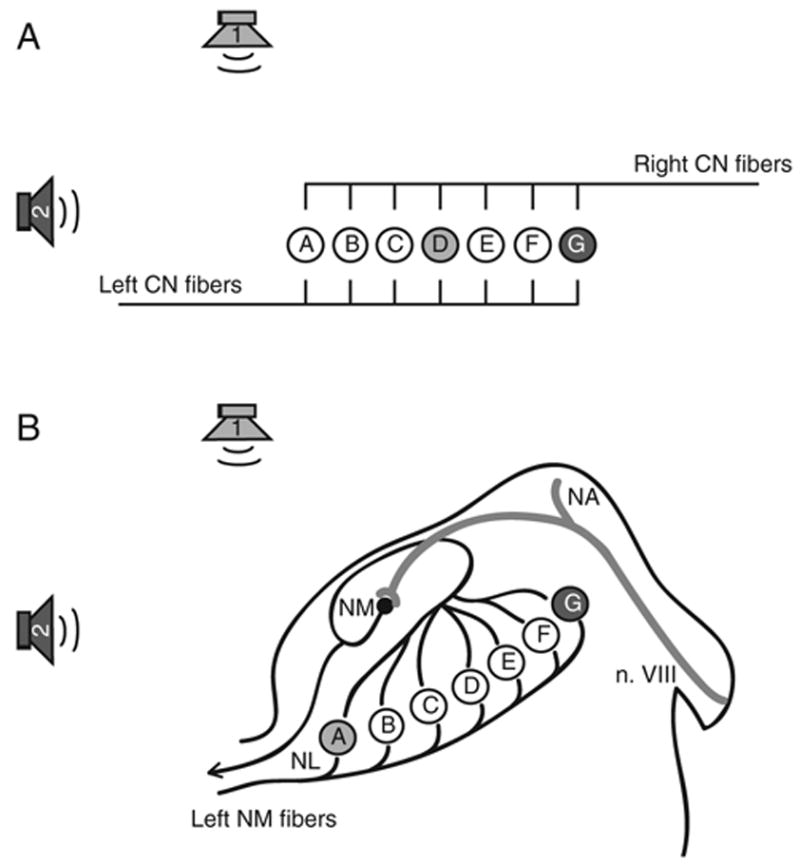
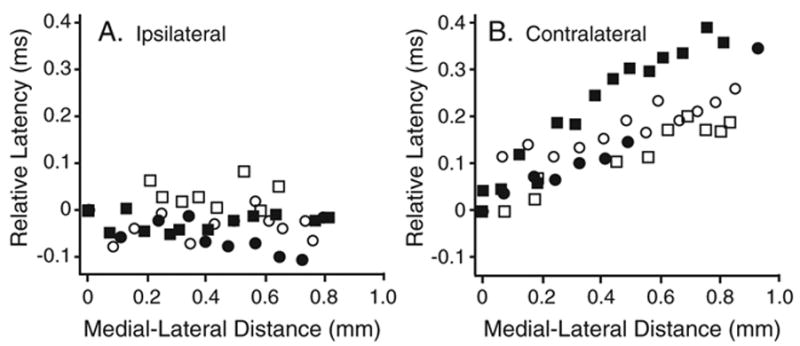
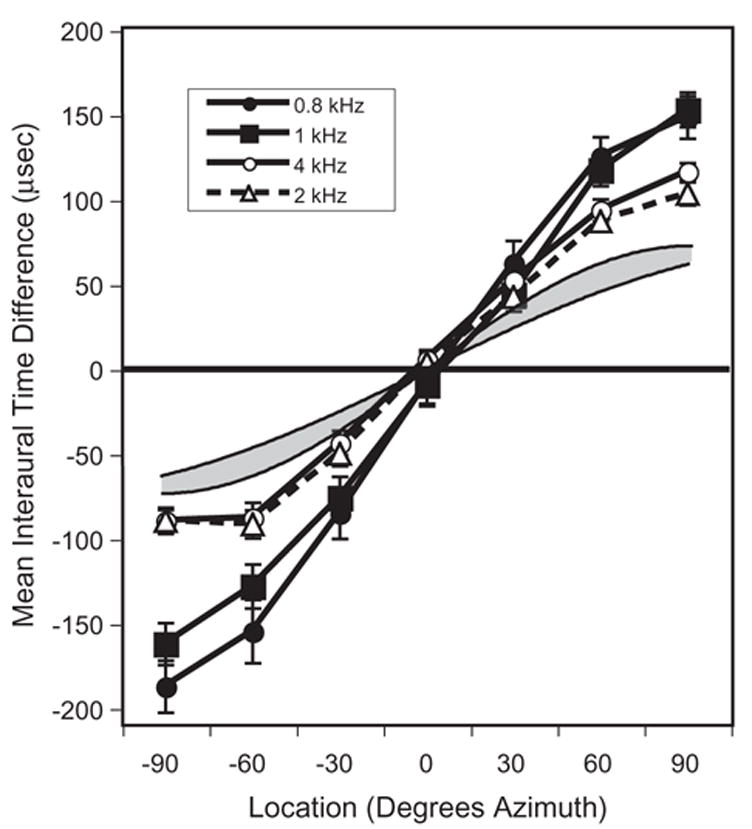
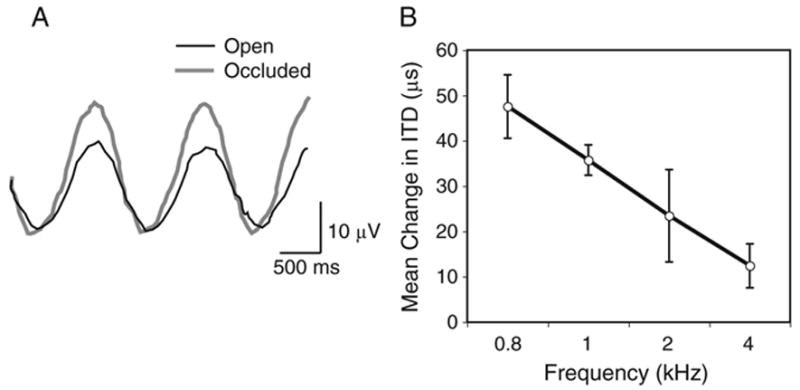
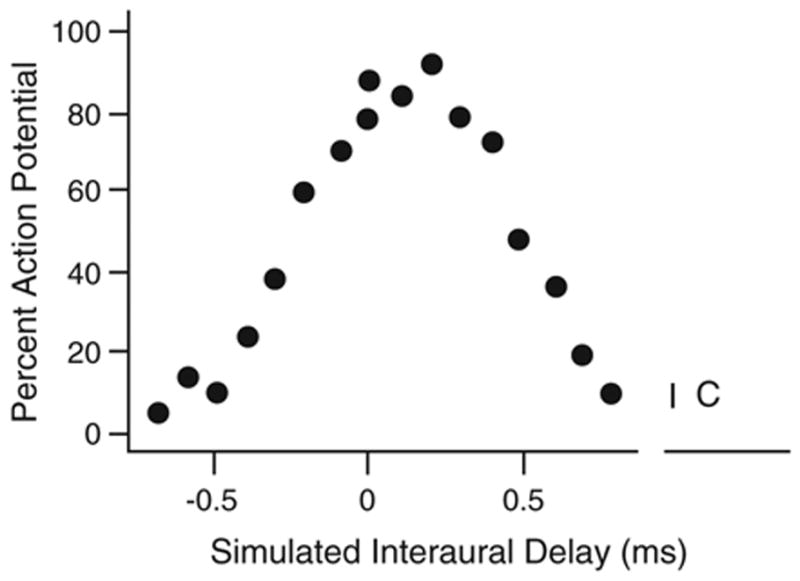
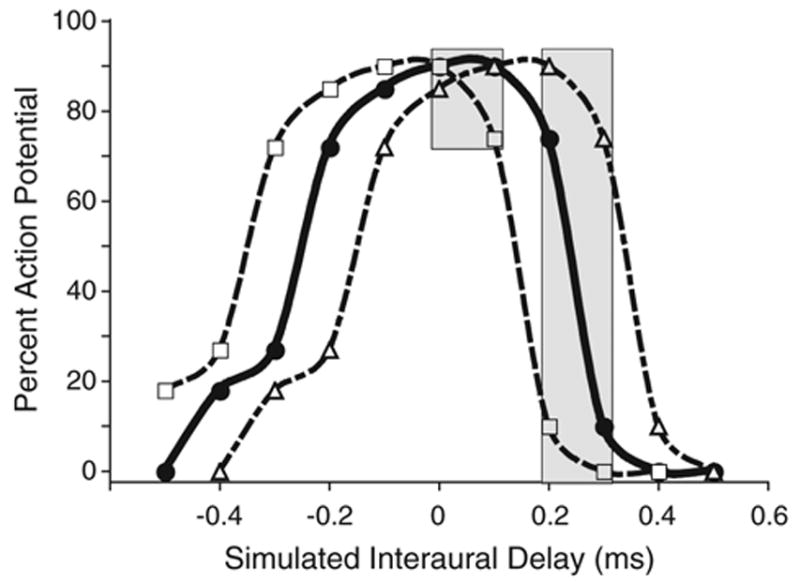
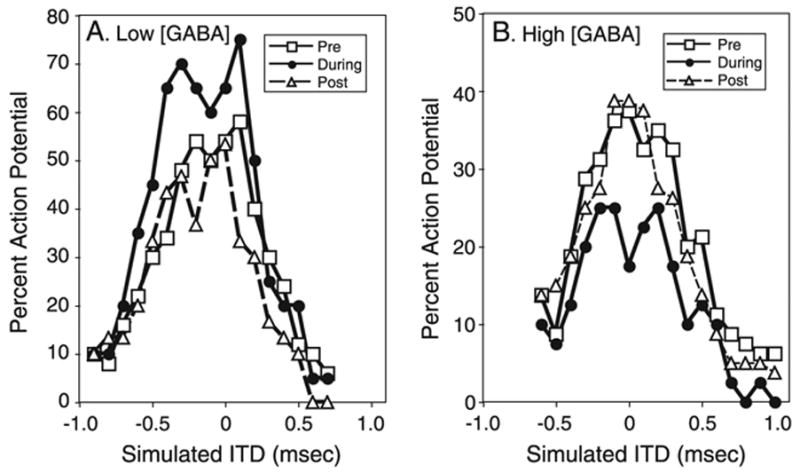
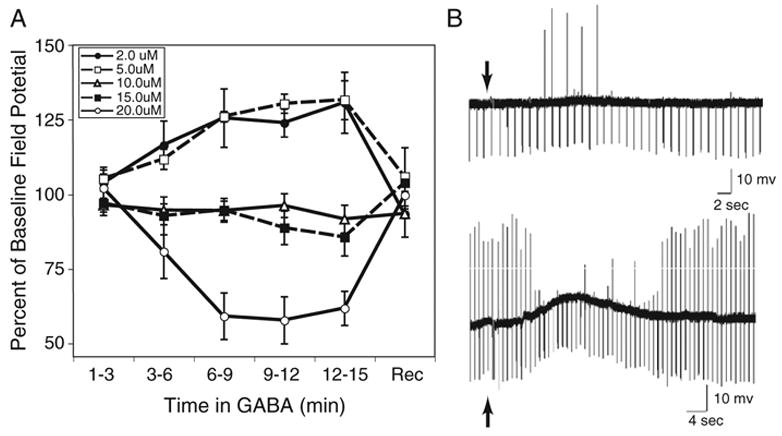
The brain stem auditory system of the chick has proven to be a useful model system for analyzing how the brain encodes temporal information. This paper reviews some of the work on a circuit in the brain stem that compares the timing of information coming from the two ears to determine the location of a sound source. The contralateral projection from the cochlear nucleus, nucleus magnocellularis (NM), to nucleus laminaris (NL) forms a delay line as it proceeds from medial to lateral across NL. NL neurons function like coincidence detectors in that they respond maximally when input from the two ears arrive simultaneously. This arrangement may allow NL to code sound space by the relative level of activity across the nucleus. The head anatomy of the chick allows for enhancement of the functional interaural time differences. Comparing the functional interaural time differences to the length of the neural delay line suggests that each NL can encode approximately one hemifield of sound space. Finally it is suggested that inhibitory input into the NM-NL circuit may provide a means to dynamically adjust the gain of the circuit to allow accurate coding of sound location despite changes in overall sound intensity.
Figures
Similar articles
-
A circuit for coding interaural time differences in the chick brainstem.J Neurosci. 1992 May;12(5):1698-708. doi: 10.1523/JNEUROSCI.12-05-01698.1992. J Neurosci. 1992. PMID: 1578264 Free PMC article.
-
Effect of GABA on the processing of interaural time differences in nucleus laminaris neurons in the chick.Eur J Neurosci. 1998 Nov;10(11):3438-50. doi: 10.1046/j.1460-9568.1998.00353.x. Eur J Neurosci. 1998. PMID: 9824457
-
Coincidence detection by binaural neurons in the chick brain stem.J Neurophysiol. 1993 Apr;69(4):1197-211. doi: 10.1152/jn.1993.69.4.1197. J Neurophysiol. 1993. PMID: 8492159
-
Creating a sense of auditory space.J Physiol. 2005 Jul 1;566(Pt 1):21-8. doi: 10.1113/jphysiol.2005.083113. Epub 2005 Mar 10. J Physiol. 2005. PMID: 15760940 Free PMC article. Review.
-
Neuronal specializations for the processing of interaural difference cues in the chick.Front Neural Circuits. 2014 May 9;8:47. doi: 10.3389/fncir.2014.00047. eCollection 2014. Front Neural Circuits. 2014. PMID: 24847212 Free PMC article. Review.
Cited by
-
Transgenic quail as a model for research in the avian nervous system: a comparative study of the auditory brainstem.J Comp Neurol. 2013 Jan 1;521(1):5-23. doi: 10.1002/cne.23187. J Comp Neurol. 2013. PMID: 22806400 Free PMC article.
-
Short-term synaptic plasticity and intensity coding.Hear Res. 2011 Sep;279(1-2):13-21. doi: 10.1016/j.heares.2011.03.001. Epub 2011 Mar 21. Hear Res. 2011. PMID: 21397676 Free PMC article. Review.
-
Glial Cell Contributions to Auditory Brainstem Development.Front Neural Circuits. 2016 Oct 21;10:83. doi: 10.3389/fncir.2016.00083. eCollection 2016. Front Neural Circuits. 2016. PMID: 27818624 Free PMC article. Review.
-
Change in the coding of interaural time difference along the tonotopic axis of the chicken nucleus laminaris.Front Neural Circuits. 2015 Aug 20;9:43. doi: 10.3389/fncir.2015.00043. eCollection 2015. Front Neural Circuits. 2015. PMID: 26347616 Free PMC article.
-
Regulation of glutamatergic and GABAergic neurotransmission in the chick nucleus laminaris: role of N-type calcium channels.Neuroscience. 2009 Dec 15;164(3):1009-19. doi: 10.1016/j.neuroscience.2009.09.013. Epub 2009 Sep 12. Neuroscience. 2009. PMID: 19751802 Free PMC article.
References
-
- Jeffress LA. A place theory of sound localization. J Comp Physiol Psychol. 1948;41:35–9. - PubMed
-
- Calford M, Piddington R. Avian interaural canal enhances interaural delay. J Comp Physiol A. 1988;162:503–10.
-
- Hill KG, Lewis DB, Hutchings ME, Coles RB. Directional hearing in the Japanese quail (Coturnix–Coturnix–japonica): I. Acoustic properties of the auditory system. J Exp Biol. 1980;86:135–51.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
