

TIM-2 is expressed on B cells and in liver and kidney and is a receptor for H-ferritin endocytosis
- PMID: 16203866
- PMCID: PMC2213179
- DOI: 10.1084/jem.20042433
TIM-2 is expressed on B cells and in liver and kidney and is a receptor for H-ferritin endocytosis
Abstract
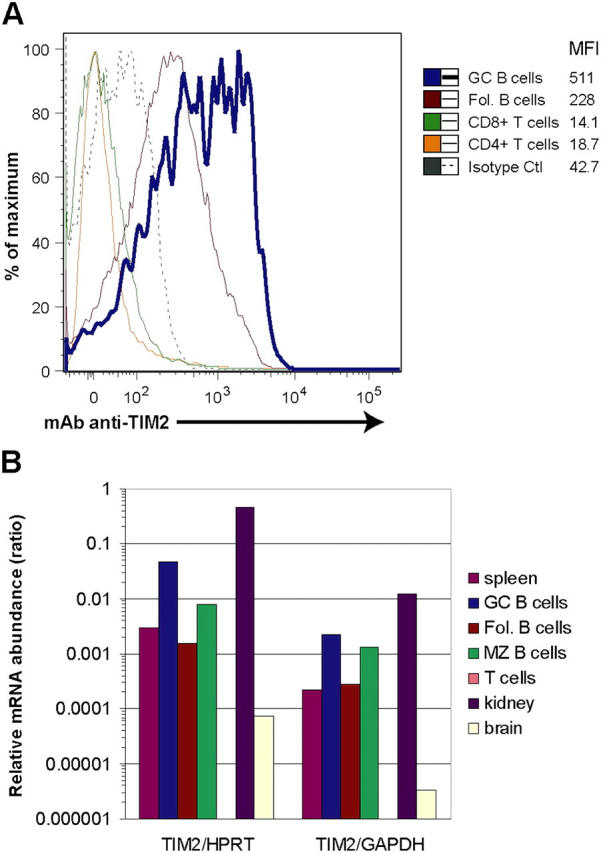
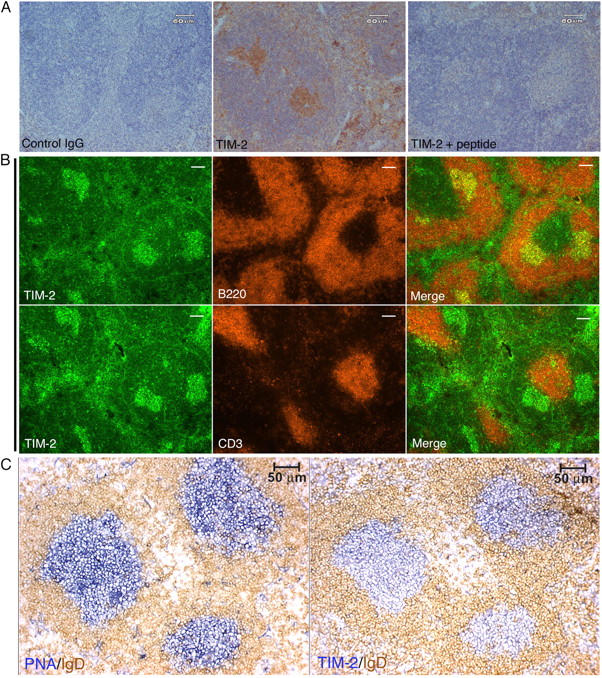
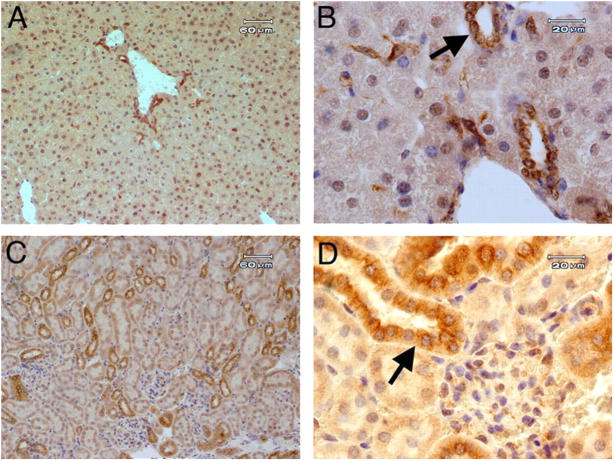
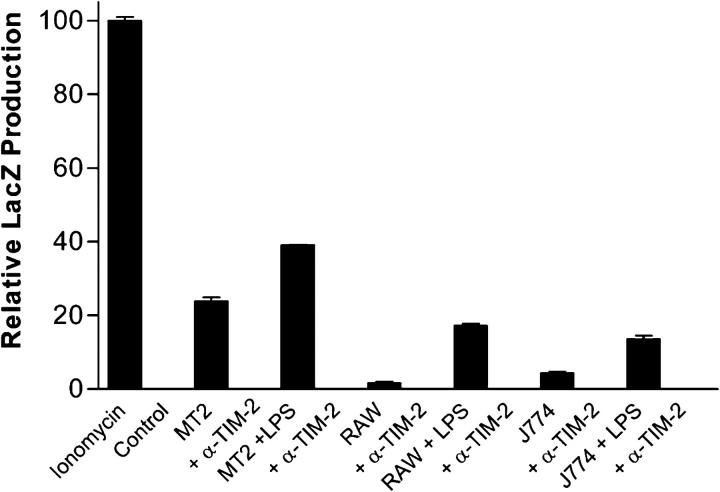
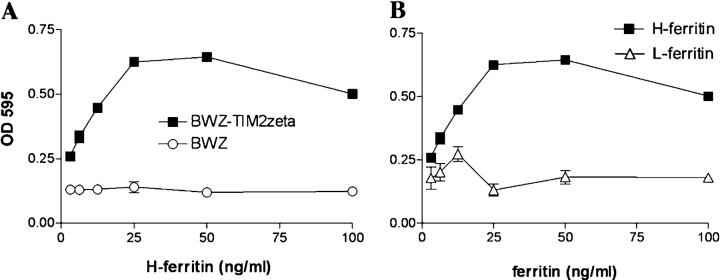
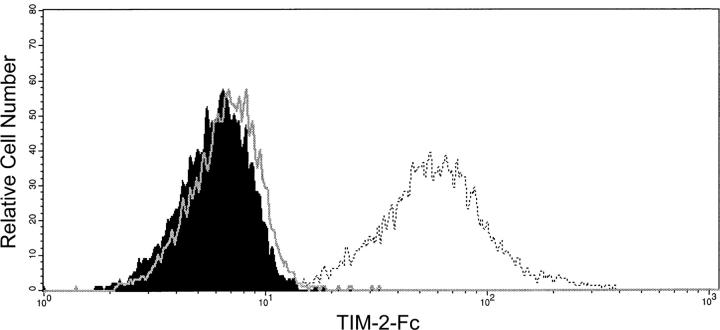
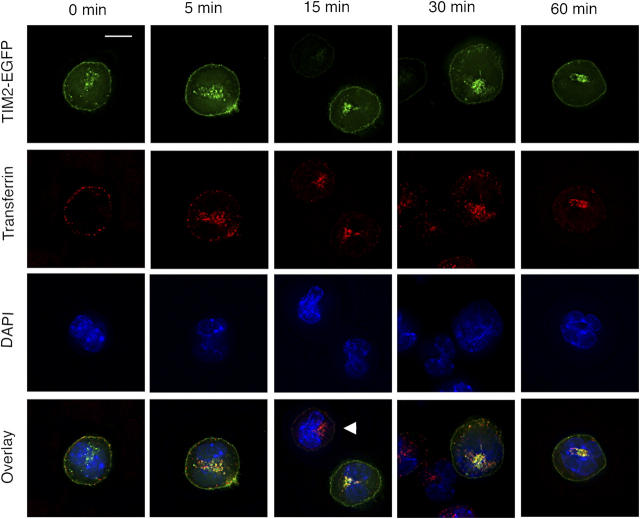
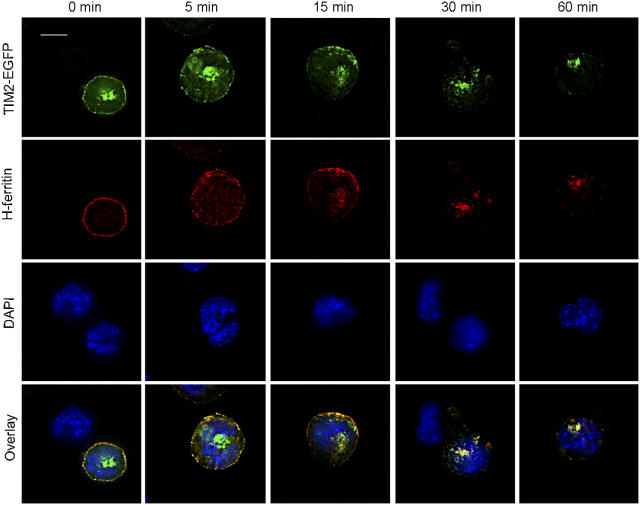
T cell immunoglobulin-domain and mucin-domain (TIM) proteins constitute a receptor family that was identified first on kidney and liver cells; recently it was also shown to be expressed on T cells. TIM-1 and -3 receptors denote different subsets of T cells and have distinct regulatory effects on T cell function. Ferritin is a spherical protein complex that is formed by 24 subunits of H- and L-ferritin. Ferritin stores iron atoms intracellularly, but it also circulates. H-ferritin, but not L-ferritin, shows saturable binding to subsets of human T and B cells, and its expression is increased in response to inflammation. We demonstrate that mouse TIM-2 is expressed on all splenic B cells, with increased levels on germinal center B cells. TIM-2 also is expressed in the liver, especially in bile duct epithelial cells, and in renal tubule cells. We further demonstrate that TIM-2 is a receptor for H-ferritin, but not for L-ferritin, and expression of TIM-2 permits the cellular uptake of H-ferritin into endosomes. This is the first identification of a receptor for ferritin and reveals a new role for TIM-2.
Figures
References
-
- Ichimura, T., J.V. Bonventre, V. Bailly, H. Wei, C.A. Hession, R.L. Cate, and M. Sanicola. 1998. Kidney injury molecule-1 (KIM-1), a putative epithelial cell adhesion molecule containing a novel immunoglobulin domain, is up-regulated in renal cells after injury. J. Biol. Chem. 273:4135–4142. - PubMed
-
- Ichimura, T., C.C. Hung, S.A. Yang, J.L. Stevens, and J.V. Bonventre. 2004. Kidney injury molecule-1: a tissue and urinary biomarker for nephrotoxicant-induced renal injury. Am. J. Physiol. Renal Physiol. 286:F552–563. - PubMed
-
- Kuchroo, V.K., D.T. Umetsu, R.H. DeKruyff, and G.J. Freeman. 2003. The TIM gene family: emerging roles in immunity and disease. Nat. Rev. Immunol. 3:454–462. - PubMed
-
- Khademi, M., Z. Illes, A.W. Gielen, M. Marta, N. Takazawa, C. Baecher-Allan, L. Brundin, J. Hannerz, C. Martin, R.A. Harris, et al. 2004. T Cell Ig- and mucin-domain-containing molecule-3 (TIM-3) and TIM-1 molecules are differentially expressed on human Th1 and Th2 cells and in cerebrospinal fluid-derived mononuclear cells in multiple sclerosis. J. Immunol. 172:7169–7176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
