

Natural killer cell and macrophage cooperation in MyD88-dependent innate responses to Plasmodium falciparum
- PMID: 16203971
- PMCID: PMC1253601
- DOI: 10.1073/pnas.0507355102
Natural killer cell and macrophage cooperation in MyD88-dependent innate responses to Plasmodium falciparum
Abstract
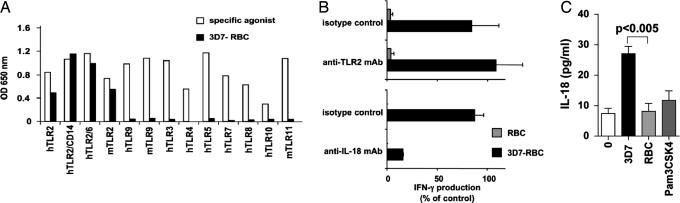
IFN-gamma secretion by natural killer (NK) cells is pivotal to several tumor and viral immune responses, during which NK and dendritic cells cooperation is required. We show here that macrophages are mandatory for NK cell IFN-gamma secretion in response to erythrocytes infected with Plasmodium falciparum (Pf), a causative agent of human malaria. In addition, direct sensing of Pf infection by NK cells induces their production of the proinflammatory chemokine CXCL8, without triggering their granule-mediated cytolytic programs. Despite their reported role in Pf recognition, Toll-like receptor (TLR) 2, TLR9, and TLR11 are individually dispensable for NK cell activation induced by Pf-infected erythrocytes. However, IL-18R expression on NK cells, IL-18 production by macrophages, and MyD88 on both cell types are essential components of this previously undescribed pathway of NK cell activation in response to a parasite infection.
Figures




References
-
- Dokun, A. O., Chu, D. T., Yang, L., Bendelac, A. S. & Yokoyama, W. M. (2001) J. Immunol. 167, 5286-5293. - PubMed
-
- Walzer, T., Dalod, M., Robbins, S. H., Zitvogel, L. & Vivier, E. (2005) Blood 106, 2252-2258. - PubMed
-
- Raulet, D. H. (2004) Nat. Immunol. 5, 996-1002. - PubMed
-
- Degli-Esposti, M. A. & Smyth, M. J. (2005) Nat. Rev. Immunol. 5, 112-124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
