

A novel murine gene, Sickle tail, linked to the Danforth's short tail locus, is required for normal development of the intervertebral disc
- PMID: 16204209
- PMCID: PMC1456172
- DOI: 10.1534/genetics.105.048934
A novel murine gene, Sickle tail, linked to the Danforth's short tail locus, is required for normal development of the intervertebral disc
Abstract
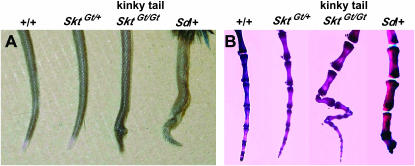
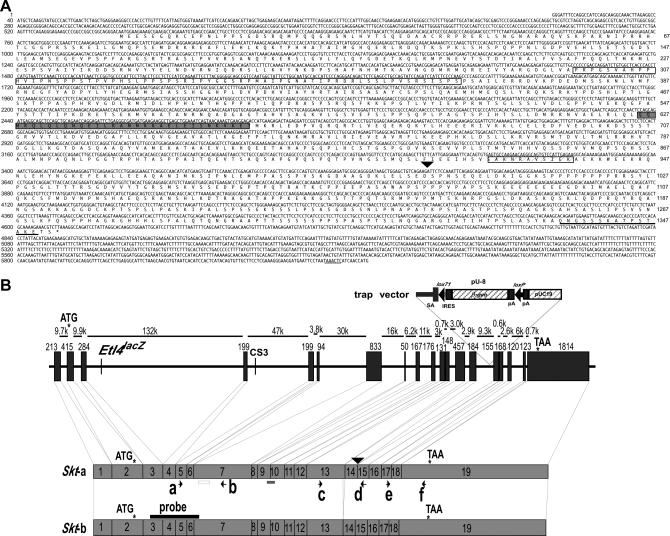
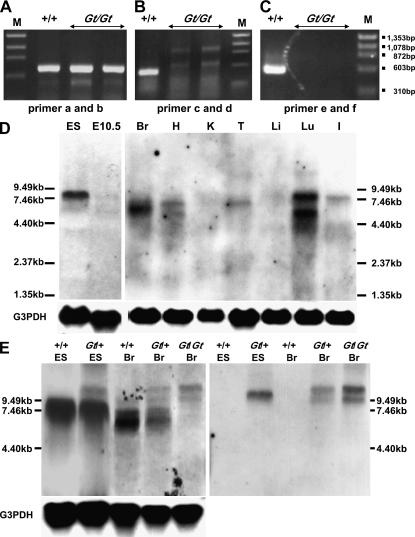
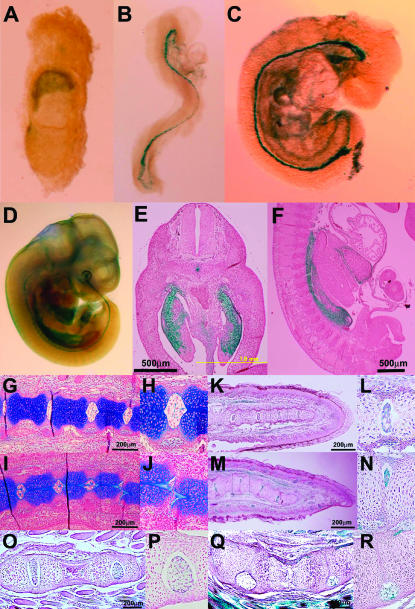
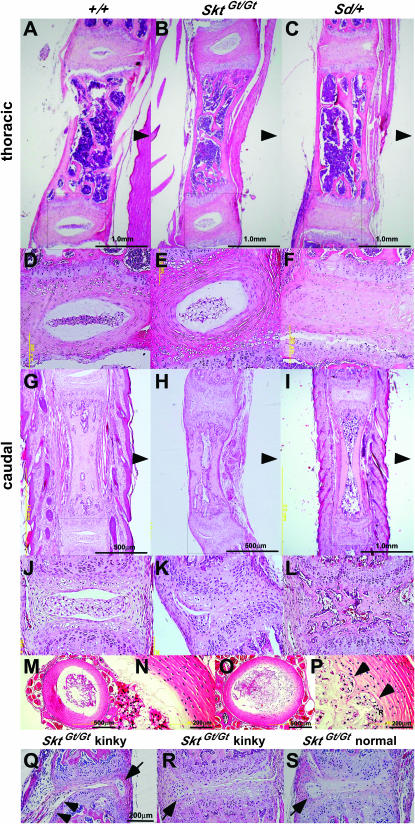
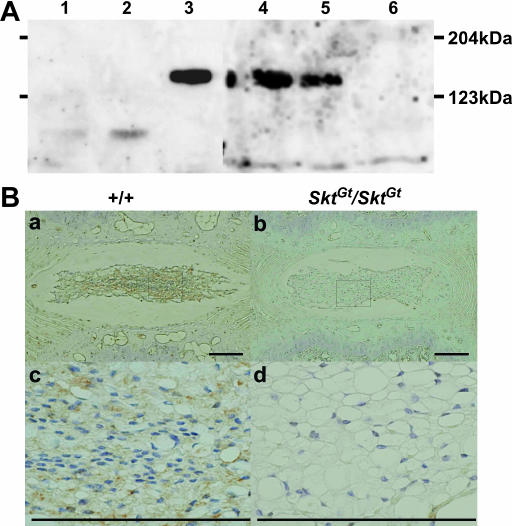
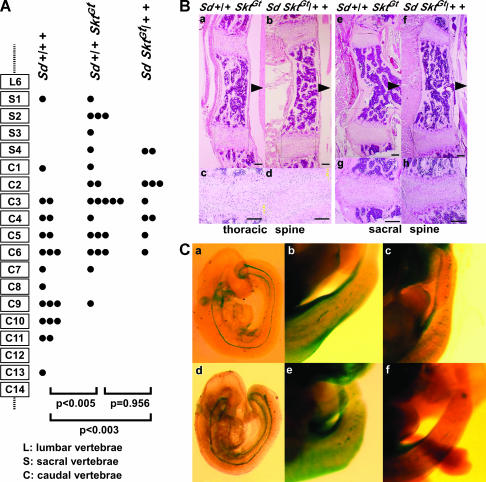
We established the mutant mouse line, B6;CB-SktGtAyu8021IMEG (SktGt), through gene-trap mutagenesis in embryonic stem cells. The novel gene identified, called Sickle tail (Skt), is composed of 19 exons and encodes a protein of 1352 amino acids. Expression of a reporter gene was detected in the notochord during embryogenesis and in the nucleus pulposus of mice. Compression of some of the nuclei pulposi in the intervertebral discs (IVDs) appeared at embryonic day (E) 17.5, resulting in a kinky-tail phenotype showing defects in the nucleus pulposus and annulus fibrosus of IVDs in SktGt/Gt mice. These phenotypes were different from those in Danforth's short tail (Sd) mice in which the nucleus pulposus was totally absent and replaced by peripheral fibers similar to those seen in the annulus fibrosus in all IVDs. The Skt gene maps to the proximal part of mouse chromosome 2, near the Sd locus. The genetic distance between them was 0.95 cM. The number of vertebrae in both [Sd +/+ SktGt] and [Sd SktGt/+ +] compound heterozygotes was less than that of Sd heterozygotes. Furthermore, the enhancer trap locus Etl4lacZ, which was previously reported to be an allele of Sd, was located in the third intron of the Skt gene.
Figures
Similar articles
-
Genetic interactions suggest that Danforth's short tail (Sd) is a gain-of-function mutation.Dev Genet. 1998;23(1):86-96. doi: 10.1002/(SICI)1520-6408(1998)23:1<86::AID-DVG9>3.0.CO;2-2. Dev Genet. 1998. PMID: 9706697
-
A Cre knock-in mouse line on the Sickle tail locus induces recombination in the notochord and intervertebral disks.Genesis. 2012 Oct;50(10):758-65. doi: 10.1002/dvg.22035. Epub 2012 May 14. Genesis. 2012. PMID: 22522943
-
The floor plate is sufficient for development of the sclerotome and spine without the notochord.Mech Dev. 2011 Jan-Feb;128(1-2):129-40. doi: 10.1016/j.mod.2010.11.005. Epub 2010 Nov 25. Mech Dev. 2011. PMID: 21111815
-
Notochord to Nucleus Pulposus Transition.Curr Osteoporos Rep. 2015 Oct;13(5):336-41. doi: 10.1007/s11914-015-0284-x. Curr Osteoporos Rep. 2015. PMID: 26231139 Review.
-
The genetics of skeletal development.Ann Genet. 1993;36(1):56-62. Ann Genet. 1993. PMID: 8503651 Review.
Cited by
-
Development, Pathogenesis, and Regeneration of the Intervertebral Disc: Current and Future Insights Spanning Traditional to Omics Methods.Front Cell Dev Biol. 2022 Mar 11;10:841831. doi: 10.3389/fcell.2022.841831. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35359439 Free PMC article. Review.
-
Genome-wide association studies of adolescent idiopathic scoliosis suggest candidate susceptibility genes.Hum Mol Genet. 2011 Apr 1;20(7):1456-66. doi: 10.1093/hmg/ddq571. Epub 2011 Jan 7. Hum Mol Genet. 2011. PMID: 21216876 Free PMC article.
-
Matrix remodeling during intervertebral disc growth and degeneration detected by multichromatic FAST staining.J Histochem Cytochem. 2009 Mar;57(3):249-56. doi: 10.1369/jhc.2008.952184. Epub 2008 Nov 11. J Histochem Cytochem. 2009. PMID: 19001641 Free PMC article.
-
A common variant rs2054564 in ADAMST17 is associated with susceptibility to lumbar spondylosis.Sci Rep. 2023 Mar 25;13(1):4900. doi: 10.1038/s41598-023-32155-w. Sci Rep. 2023. PMID: 36966180 Free PMC article.
-
Copy number variants suggest different molecular pathways for the pathogenesis of bladder exstrophy.Am J Med Genet A. 2023 Feb;191(2):378-390. doi: 10.1002/ajmg.a.63031. Epub 2022 Nov 8. Am J Med Genet A. 2023. PMID: 36349425 Free PMC article.
References
-
- Alfred, J. B., K. Rance, B. A. Taylor, S. J. Phillips, C. M. Abbott et al., 1997. Mapping in the region of Danforth's short tail and the localization of tail length modifiers. Genome Res. 7: 108–117. - PubMed
-
- Allen, N. D., D. G. Cran, S. C. Barton, S. Hettel, W. Reik et al., 1988. Transgenes as probes for active chromosomal domains in mouse development. Nature 333: 852–855. - PubMed
-
- Ang, S., and J. Rossant, 1994. HNF-3β is essential for node and notochord formation in mouse development. Cell 78: 561–574. - PubMed
-
- Araki, K., T. Imaizumi, T. Sekimoto, K. Yoshinobu, J. Yoshimuta et al., 1999. Exchangeable gene trap using the Cre/mutated lox system. Cell. Mol. Biol. 45: 737–750. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
