

A catalog of stability-associated sequence elements in 3' UTRs of yeast mRNAs
- PMID: 16207357
- PMCID: PMC1257469
- DOI: 10.1186/gb-2005-6-10-r86
A catalog of stability-associated sequence elements in 3' UTRs of yeast mRNAs
Abstract
Background: In recent years, intensive computational efforts have been directed towards the discovery of promoter motifs that correlate with mRNA expression profiles. Nevertheless, it is still not always possible to predict steady-state mRNA expression levels based on promoter signals alone, suggesting that other factors may be involved. Other genic regions, in particular 3' UTRs, which are known to exert regulatory effects especially through controlling RNA stability and localization, were less comprehensively investigated, and deciphering regulatory motifs within them is thus crucial.
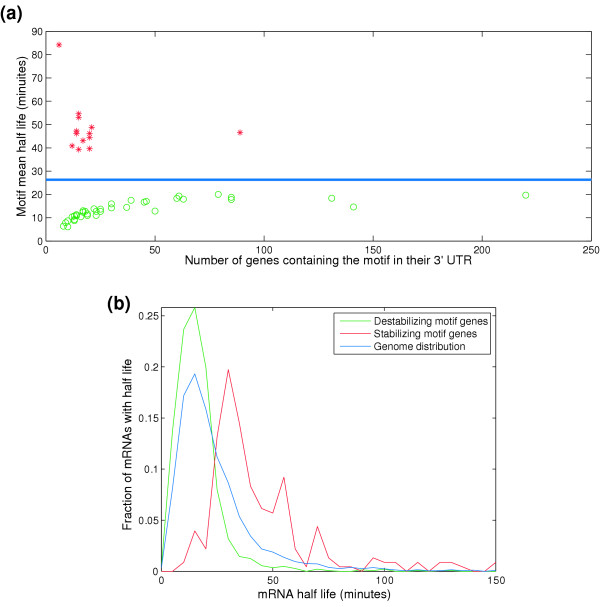
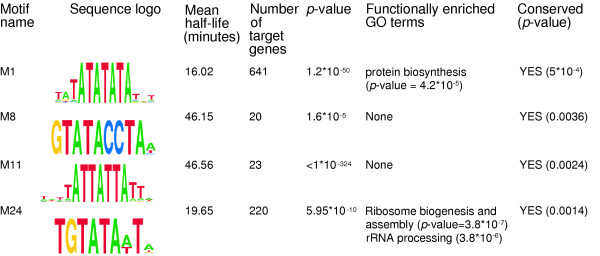
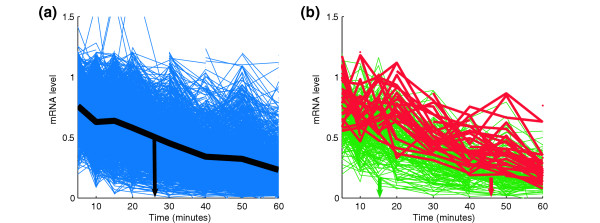
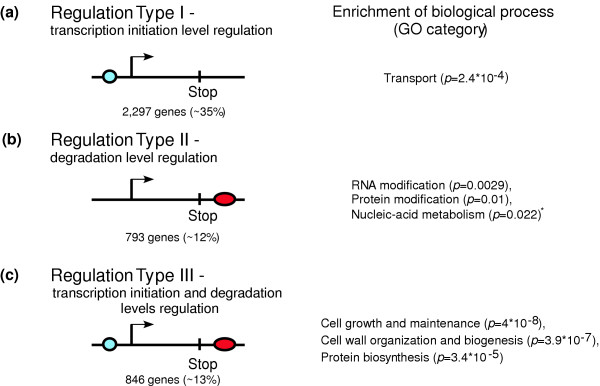
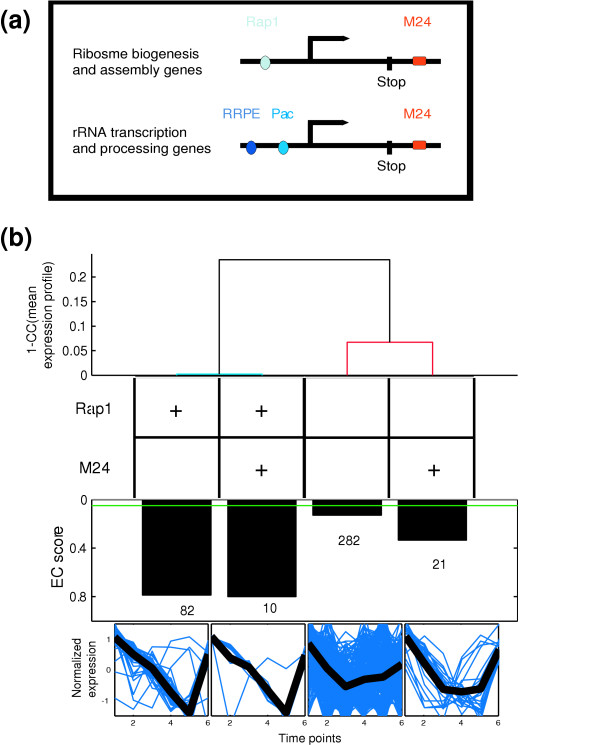
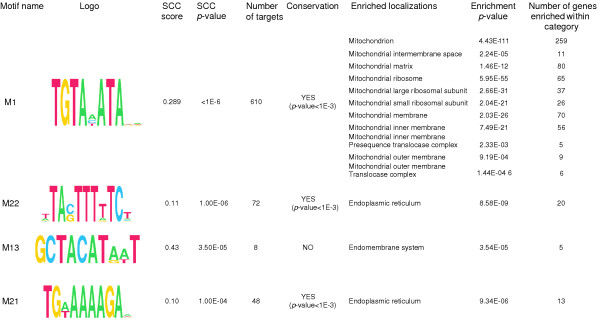
Results: By analyzing 3' UTR sequences and mRNA decay profiles of Saccharomyces cerevisiae genes, we derived a catalog of 53 sequence motifs that may be implicated in stabilization or destabilization of mRNAs. Some of the motifs correspond to known RNA-binding protein sites, and one of them may act in destabilization of ribosome biogenesis genes during stress response. In addition, we present for the first time a catalog of 23 motifs associated with subcellular localization. A significant proportion of the 3' UTR motifs is highly conserved in orthologous yeast genes, and some of the motifs are strikingly similar to recently published mammalian 3' UTR motifs. We classified all genes into those regulated only at transcription initiation level, only at degradation level, and those regulated by a combination of both. Interestingly, different biological functionalities and expression patterns correspond to such classification.
Conclusion: The present motif catalogs are a first step towards the understanding of the regulation of mRNA degradation and subcellular localization, two important processes which--together with transcription regulation--determine the cell transcriptome.
Figures

References
-
- Segal E, Shapira M, Regev A, Pe'er D, Botstein D, Koller D, Friedman N. Module networks: identifying regulatory modules and their condition-specific regulators from gene expression data. Nat Genet . 2003;34:166–176. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
