

Differential regulation of cyclooxygenase-2 in nontransformed and ras-transformed intestinal epithelial cells
- PMID: 16207478
- PMCID: PMC1501890
- DOI: 10.1593/neo.04652
Differential regulation of cyclooxygenase-2 in nontransformed and ras-transformed intestinal epithelial cells
Abstract
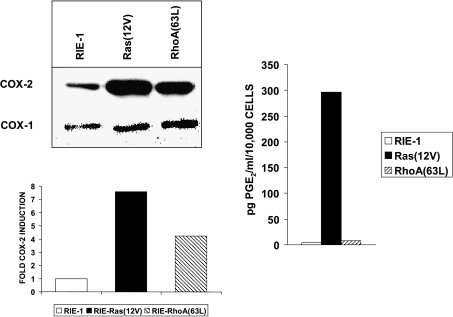
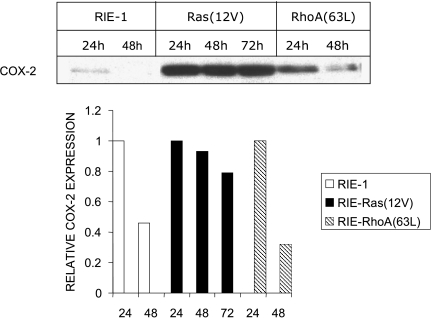
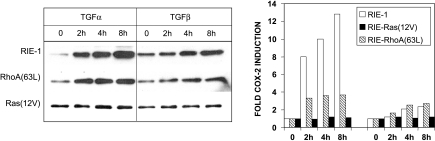
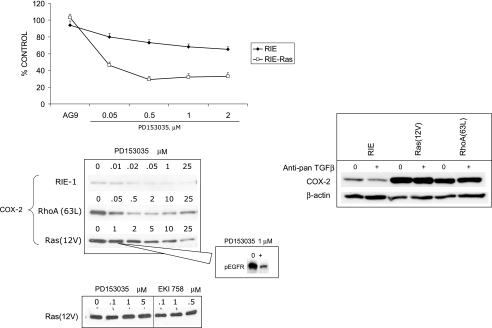
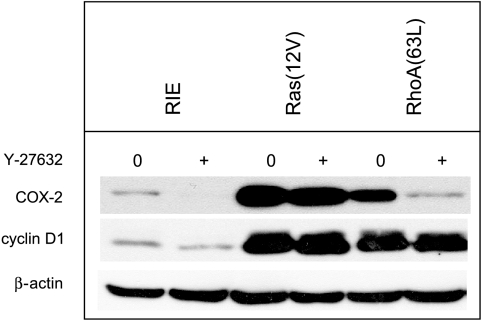
To determine signaling pathways responsible for modulation of COX-2 expression in nontransformed and transformed epithelial cells, we studied a rat intestinal epithelial (RIE) cell line expressing constitutively active Ras and RhoA. Expression of COX-2 protein was higher in RIE-RhoA(63L) (four-fold) and RIE-Ras(12V) (seven-fold) cells than in parental cells. Prior work suggests that Ras hyperactivity induces the expression of transforming growth factor (TGF)beta and increases epidermal growth factor (EGF)-related peptide signaling-possible mechanisms for increased COX-2 expression. Expression of COX-2 was stimulated by TGFbeta and TGFalpha in RIE and RIE-Rho(63L) cells, but not further stimulated in RIE-Ras(12V) cells. PD153035, an inhibitor of EGF receptor tyrosine kinase, and PD98059, an inhibitor of Erk, attenuated COX-2 expression in RIE and RIE-RhoA(63L). However, the high levels of COX-2 expression in RIE-Ras(12V) cells were not inhibited by either compound. Titration with a pan-neutralizing anti-TGFbeta antibody did not decrease COX-2 in RIE-Ras(12V) cells, even with concurrent EGFR inhibition. Thus, stimulation of the EGF receptor is important in the modulation of COX-2 expression in nontransformed RIE and RIE-RhoA(63L) cells. In Ras-transformed cells, signaling by additional Ras effector pathways, perhaps the RhoA pathway, must be invoked. Identification of these pathways is critical for therapeutic manipulation of COX-2 expression.
Figures

Similar articles
-
Raf and RhoA cooperate to transform intestinal epithelial cells and induce growth resistance to transforming growth factor beta.Mol Cancer Res. 2004 Apr;2(4):233-41. Mol Cancer Res. 2004. PMID: 15140945
-
A raf-independent epidermal growth factor receptor autocrine loop is necessary for Ras transformation of rat intestinal epithelial cells.J Biol Chem. 1997 Jul 25;272(30):18926-31. doi: 10.1074/jbc.272.30.18926. J Biol Chem. 1997. PMID: 9228072
-
Transformation by oncogenic Ras expands the early genomic response to transforming growth factor beta in intestinal epithelial cells.Neoplasia. 2008 Oct;10(10):1073-82. doi: 10.1593/neo.07739. Neoplasia. 2008. PMID: 18813357 Free PMC article.
-
Growth inhibitory signalling by TGFbeta is blocked in Ras-transformed intestinal epithelial cells at a post-receptor locus.Cell Signal. 2003 Jul;15(7):699-708. doi: 10.1016/s0898-6568(03)00010-x. Cell Signal. 2003. PMID: 12742230
-
Ras-mediated suppression of TGFbetaRII expression in intestinal epithelial cells involves Raf-independent signaling.Neoplasia. 2000 Jul-Aug;2(4):357-64. doi: 10.1038/sj.neo.7900099. Neoplasia. 2000. PMID: 11005570 Free PMC article.
Cited by
-
A review of the past, present, and future directions of neoplasia.Neoplasia. 2005 Dec;7(12):1039-46. doi: 10.1593/neo.05793. Neoplasia. 2005. PMID: 16354585 Free PMC article. Review. No abstract available.
-
Hyaluronan constitutively regulates activation of COX-2-mediated cell survival activity in intestinal epithelial and colon carcinoma cells.J Biol Chem. 2008 May 23;283(21):14335-44. doi: 10.1074/jbc.M703811200. Epub 2008 Mar 7. J Biol Chem. 2008. PMID: 18326857 Free PMC article.
-
Celecoxib enhances the anti-inflammatory effects of farnesylthiosalicylic acid on T cells independent of prostaglandin E(2) production.Inflammation. 2012 Oct;35(5):1706-14. doi: 10.1007/s10753-012-9488-8. Inflammation. 2012. PMID: 22688643
-
Angiotensin II bi-directionally regulates cyclooxygenase-2 expression in intestinal epithelial cells.Mol Cell Biochem. 2008 Aug;315(1-2):185-93. doi: 10.1007/s11010-008-9806-5. Epub 2008 Jun 10. Mol Cell Biochem. 2008. PMID: 18543083 Free PMC article.
-
Epidermal growth factor increases prostaglandin E2 production via ERK1/2 MAPK and NF-kappaB pathway in fibroblast like synoviocytes from patients with rheumatoid arthritis.Rheumatol Int. 2010 Feb;30(4):443-9. doi: 10.1007/s00296-009-0976-6. Epub 2009 Aug 13. Rheumatol Int. 2010. PMID: 19680656
References
-
- FitzGerald G, Patrono C. The coxibs, selective inhibitors of cyclooxygenase-2. N Engl J Med. 2001;345:433–442. - PubMed
-
- Eberhart CE, Coffey RJ, Radhik A, Giardiello FM, Ferrenbach S, DuBois RN. Up-regulation of cyclooxygenase 2 gene expression in human colorectal adenomas and adenocarcinomas. Gastroenterology. 1994;107:1183–1188. - PubMed
-
- Williams CS, Luongo C, Radhika A, Zhang T, Lamps LW, Nanney LB, Beauchamp RD, DuBois RN. Elevated cyclooxygenase-2 levels in Min mouse adenomas. Gastroenterology. 1996;111:1134–1140. - PubMed
-
- Shao J, Sheng H, Aramandla R, Pereira MA, Lubet RA, Hawk E, Grogan L, Kirsch IR, Washington MK, Beauchamp RD, et al. Coordinate regulation of cyclooxygenase-2 and TGF-beta1 in replication error-positive colon cancer and azoxymethane-induced rat colonic tumors. Carcinogenesis. 1999;20:185–191. - PubMed
-
- DuBois RN. Cyclooxygenase-2 and colorectal cancer. Prog Exp Tumor Res. 2003;37:124–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous