

Germinal center exclusion of autoreactive B cells is defective in human systemic lupus erythematosus
- PMID: 16211091
- PMCID: PMC1242189
- DOI: 10.1172/JCI24179
Germinal center exclusion of autoreactive B cells is defective in human systemic lupus erythematosus
Abstract
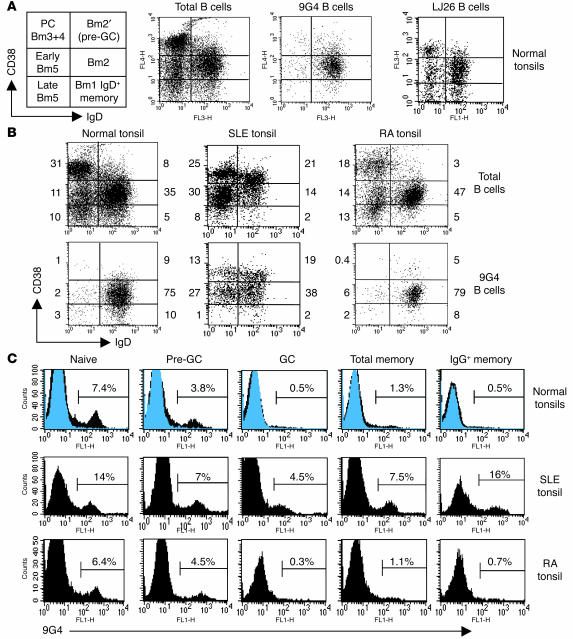
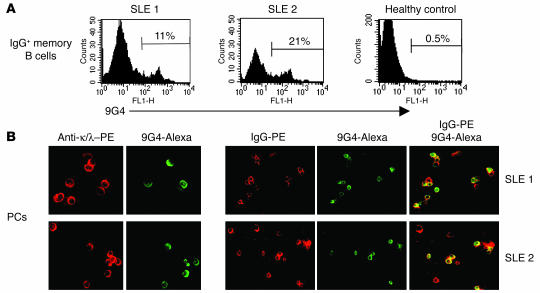
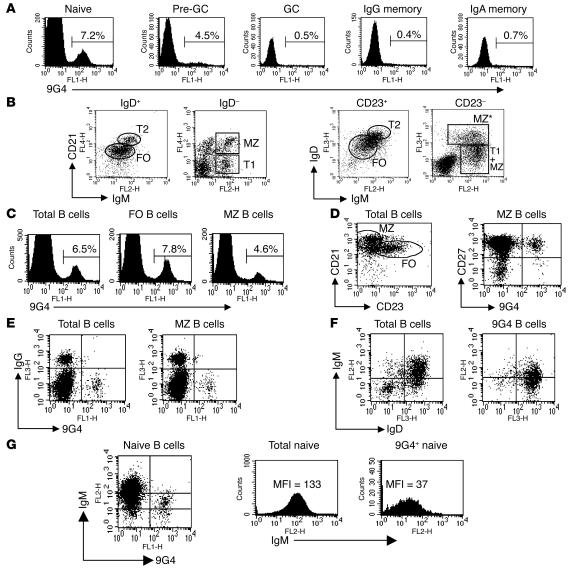
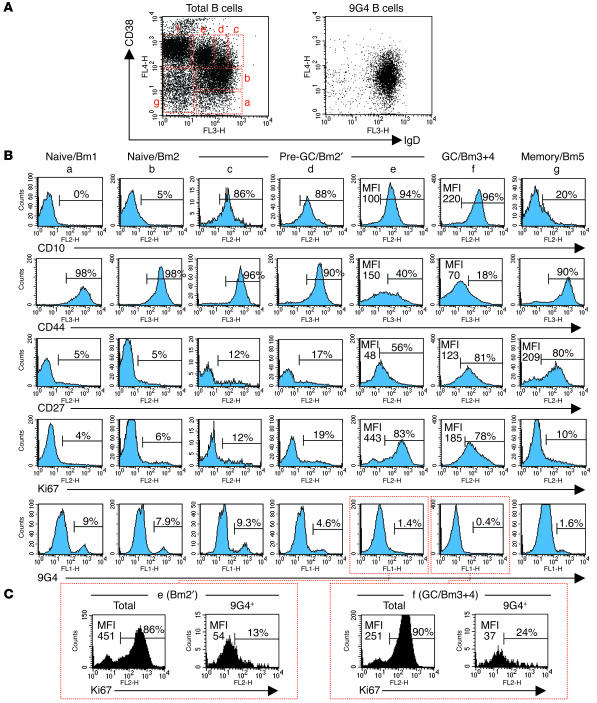
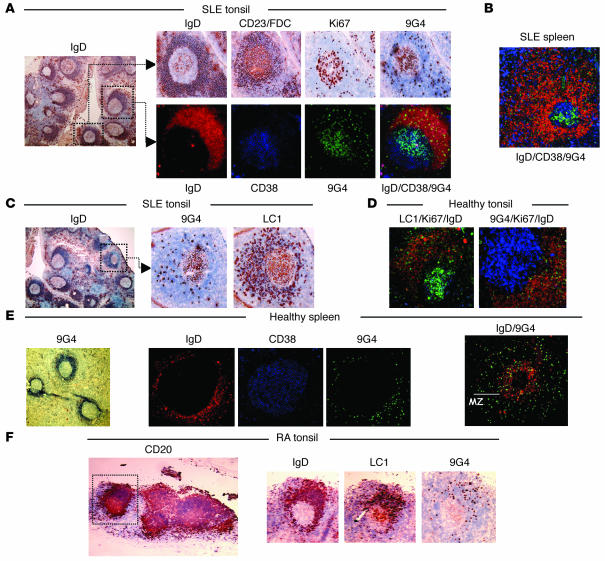
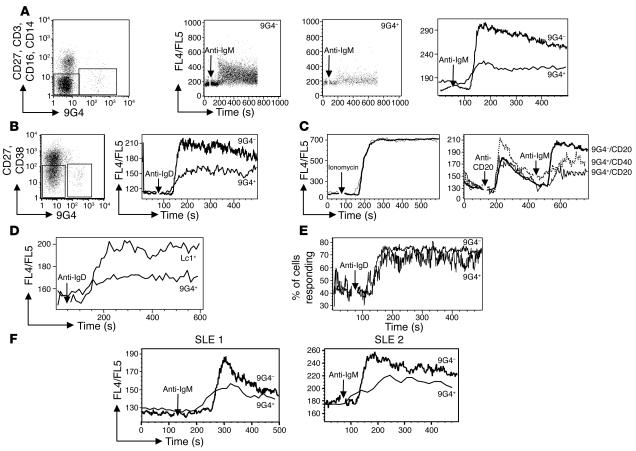
Breach of B cell tolerance is central to the pathogenesis of systemic lupus erythematosus (SLE). However, how B cell tolerance is subverted in human SLE is poorly understood due to difficulties in identifying relevant autoreactive B cells and in obtaining lymphoid tissue. We have circumvented these limitations by using tonsil biopsies to study autoreactive B cells (9G4 B cells), whose regulation is abnormal in SLE. Here we show that 9G4 B cells are physiologically excluded during the early stages of the GC reaction before acquiring a centroblast phenotype. Furthermore, we provide evidence to indicate that an anergic response to B cell receptor stimulation may be responsible for such behavior. In contrast, in SLE, 9G4 B cells progressed unimpeded through this checkpoint, successfully participated in GC reactions, and expanded within the post-GC IgG memory and plasma cell compartments. The faulty regulation of 9G4 B cells was not shared by RA patients. To our knowledge, this work represents the first comparative analysis of the fate of a specific autoreactive human B cell population. The results identify a defective tolerance checkpoint that appears to be specific for human SLE.
Figures
References
-
- Hochberg MC. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus [letter] Arthritis Rheum. 1997;40:1725. - PubMed
-
- Winfield JB, Winchester RJ, Kunkel HG. Association of cold-reactive antilymphocyte antibodies with lymphopenia in systemic lupus erythematosus. Arthritis Rheum. 1975;18:587–594. - PubMed
-
- Cappione AJ, Pugh-Bernard AE, Anolik JH, Sanz I. Lupus IgG VH4.34 antibodies bind to a 220-kDa glycoform of CD45/B220 on the surface of human B lymphocytes. J. Immunol. 2004;172:4298–4307. - PubMed
-
- Chan OT, Madaio MP, Shlomchik MJ. The central and multiple roles of B cells in lupus pathogenesis. Immunol. Rev. 1999;169:107–121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
