

Ras1-induced hyphal development in Candida albicans requires the formin Bni1
- PMID: 16215178
- PMCID: PMC1265897
- DOI: 10.1128/EC.4.10.1712-1724.2005
Ras1-induced hyphal development in Candida albicans requires the formin Bni1
Abstract
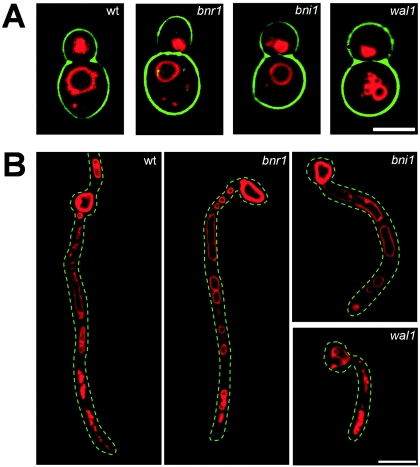
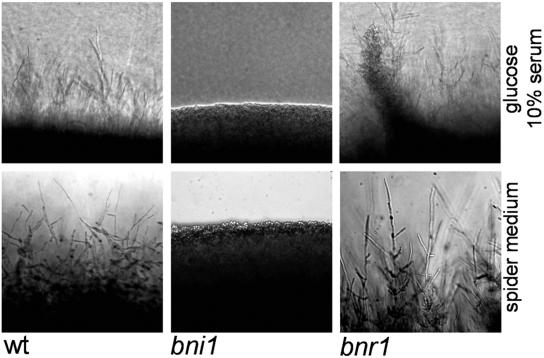
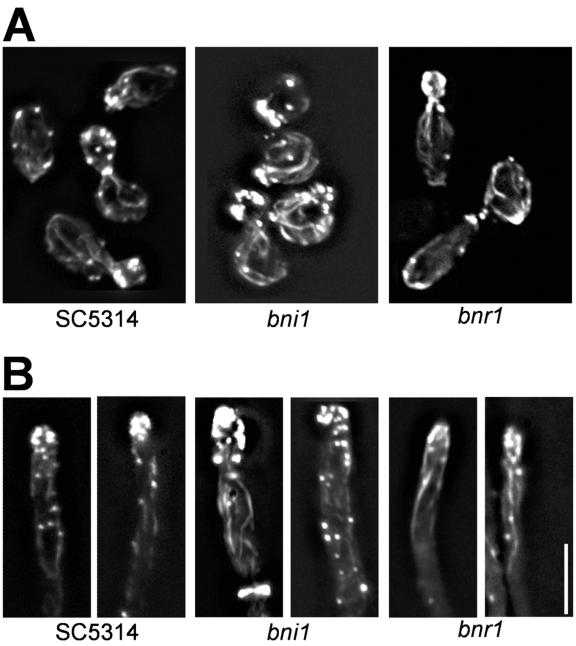
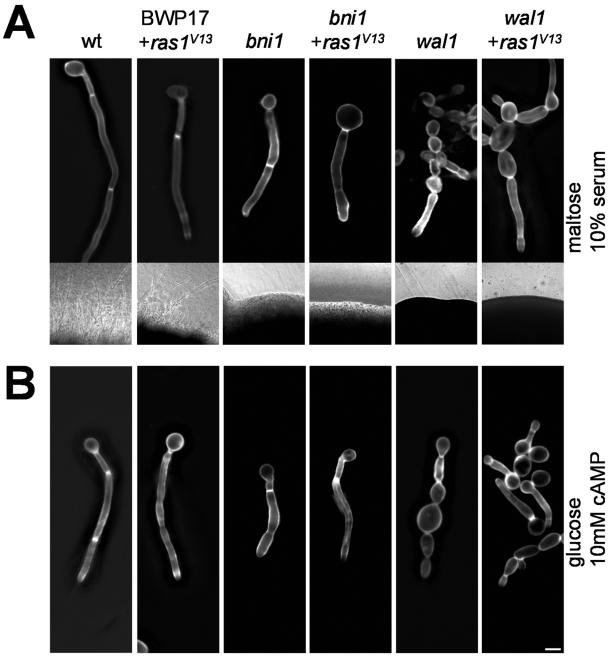
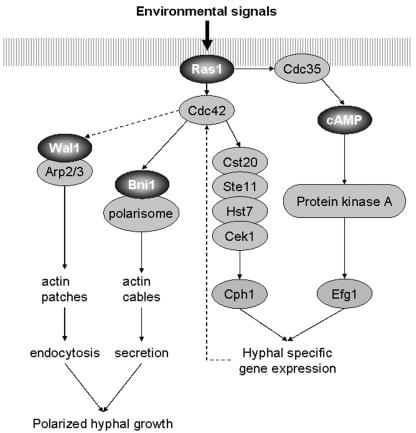
Formins are downstream effector proteins of Rho-type GTPases and are involved in the organization of the actin cytoskeleton and actin cable assembly at sites of polarized cell growth. Here we show using in vivo time-lapse microscopy that deletion of the Candida albicans formin homolog BNI1 results in polarity defects during yeast growth and hyphal stages. Deletion of the second C. albicans formin, BNR1, resulted in elongated yeast cells with cell separation defects but did not interfere with the ability of bnr1 cells to initiate and maintain polarized hyphal growth. Yeast bni1 cells were swollen, showed an increased random budding pattern, and had a severe defect in cytokinesis, with enlarged bud necks. Induction of hyphal development in bni1 cells resulted in germ tube formation but was halted at the step of polarity maintenance. Bni1-green fluorescent protein is found persistently at the hyphal tip and colocalizes with a structure resembling the Spitzenkörper of true filamentous fungi. Introduction of constitutively active ras1G13V in the bni1 strain or addition of cyclic AMP to the growth medium did not bypass bni1 hyphal growth defects. Similarly, these agents were not able to suppress hyphal growth defects in the wal1 mutant which is lacking the Wiskott-Aldrich syndrome protein (WASP) homolog. These results suggest that the maintenance of polarized hyphal growth in C. albicans requires coordinated regulation of two actin cytoskeletal pathways, including formin-mediated secretion and WASP-dependent endocytosis.
Figures



Similar articles
-
Candida albicans Rho-type GTPase-encoding genes required for polarized cell growth and cell separation.Eukaryot Cell. 2007 May;6(5):844-54. doi: 10.1128/EC.00201-06. Epub 2007 Mar 9. Eukaryot Cell. 2007. PMID: 17351079 Free PMC article.
-
The formin family protein CaBni1p has a role in cell polarity control during both yeast and hyphal growth in Candida albicans.J Cell Sci. 2005 Jun 15;118(Pt 12):2637-48. doi: 10.1242/jcs.02393. Epub 2005 May 24. J Cell Sci. 2005. PMID: 15914538
-
Role of CaBud6p in the polarized growth of Candida albicans.J Microbiol. 2006 Jun;44(3):311-9. J Microbiol. 2006. PMID: 16820761
-
Germ tube growth of Candida albicans.Curr Top Med Mycol. 1997 Dec;8(1-2):43-55. Curr Top Med Mycol. 1997. PMID: 9504066 Review.
-
Cell polarity in filamentous fungi: shaping the mold.Int Rev Cytol. 2006;251:41-77. doi: 10.1016/S0074-7696(06)51002-2. Int Rev Cytol. 2006. PMID: 16939777 Review.
Cited by
-
Regulation of Candida albicans Hyphal Morphogenesis by Endogenous Signals.J Fungi (Basel). 2019 Feb 28;5(1):21. doi: 10.3390/jof5010021. J Fungi (Basel). 2019. PMID: 30823468 Free PMC article. Review.
-
Candida albicans Rho-type GTPase-encoding genes required for polarized cell growth and cell separation.Eukaryot Cell. 2007 May;6(5):844-54. doi: 10.1128/EC.00201-06. Epub 2007 Mar 9. Eukaryot Cell. 2007. PMID: 17351079 Free PMC article.
-
Ras-Mediated Signal Transduction and Virulence in Human Pathogenic Fungi.Fungal Genom Biol. 2012;2(1):105. doi: 10.4172/2165-8056.1000105. Fungal Genom Biol. 2012. PMID: 24855584 Free PMC article.
-
A Myosin Light Chain Is Critical for Fungal Growth Robustness in Candida albicans.mBio. 2021 Oct 26;12(5):e0252821. doi: 10.1128/mBio.02528-21. Epub 2021 Oct 5. mBio. 2021. PMID: 34607458 Free PMC article.
-
Growth of Candida albicans hyphae.Nat Rev Microbiol. 2011 Aug 16;9(10):737-48. doi: 10.1038/nrmicro2636. Nat Rev Microbiol. 2011. PMID: 21844880 Review.
References
-
- Bartnicki-Garcia, S., D. D. Bartnicki, G. Gierz, R. Lopez-Franco, and C. E. Bracker. 1995. Evidence that Spitzenkörper behavior determines the shape of a fungal hypha: a test of the hyphoid model. Exp. Mycol. 19:153-159. - PubMed
-
- Bartnicki-Garcia, S., F. Hergert, and G. Gierz. 1989. Computer simulation of fungal morphogenesis and the mathematical basis for hyphal tip growth. Protoplasma 153:46-57.
-
- Berman, J., and P. E. Sudbery. 2002. Candida albicans: a molecular revolution built on lessons from budding yeast. Nat. Rev. Genet. 3:918-930. - PubMed
-
- Borkovich, K. A., L. A. Alex, O. Yarden, M. Freitag, G. E. Turner, N. D. Read, S. Seiler, D. Bell-Pedersen, J. Paietta, N. Plesofsky, M. Plamann, M. Goodrich-Tanrikulu, U. Schulte, G. Mannhaupt, F. E. Nargang, A. Radford, C. Selitrennikoff, J. E. Galagan, J. C. Dunlap, J. J. Loros, D. Catcheside, H. Inoue, R. Aramayo, M. Polymenis, E. U. Selker, M. S. Sachs, G. A. Marzluf, I. Paulsen, R. Davis, D. J. Ebbole, A. Zelter, E. R. Kalkman, R. O'Rourke, F. Bowring, J. Yeadon, C. Ishii, K. Suzuki, W. Sakai, and R. Pratt. 2004. Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68:1-108. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
