

Low mobility of phosphatidylinositol 4,5-bisphosphate underlies receptor specificity of Gq-mediated ion channel regulation in atrial myocytes
- PMID: 16217031
- PMCID: PMC1257688
- DOI: 10.1073/pnas.0408851102
Low mobility of phosphatidylinositol 4,5-bisphosphate underlies receptor specificity of Gq-mediated ion channel regulation in atrial myocytes
Abstract
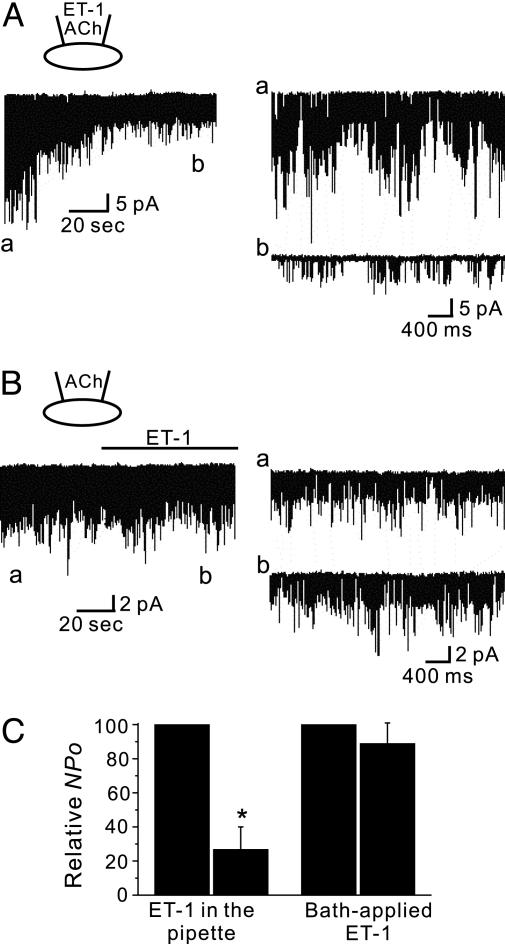
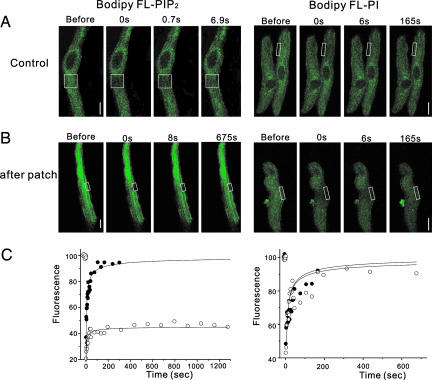
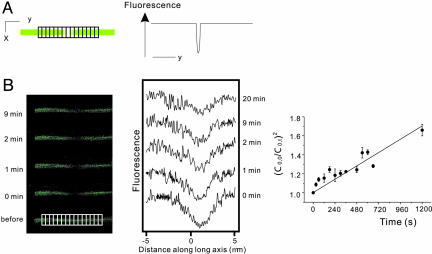
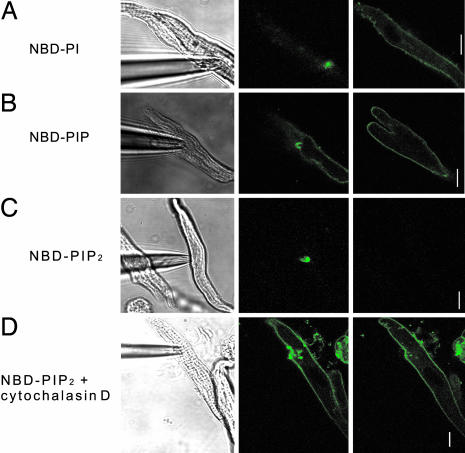
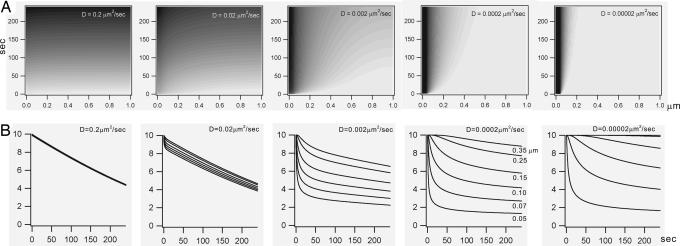
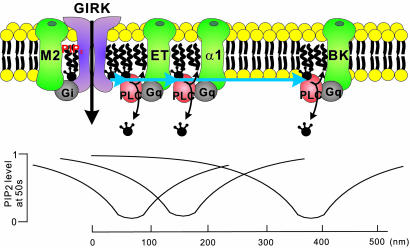
We have shown previously that cardiac G protein-gated inwardly rectifying K+ (GIRK) channels are inhibited by Gq protein-coupled receptors (GqPCRs) via phosphatidylinositol 4,5-bisphosphate (PIP2) depletion in a receptor-specific manner. To investigate the mechanism of receptor specificity, we examined whether the activation of GqPCRs induces localized PIP2 depletion. When we applied endothelin-1 to the bath, GIRK channel activities recorded in cell-attached patches were not changed, implying that PIP2 signal is not diffusible but is a localized signal. To test this possibility, we directly measured lateral diffusion by introducing fluorescence-labeled phosphoinositides to a small area of the membrane with patch pipettes. After pipettes were attached, phosphatidylinositol 4-monophosphate or phosphatidylinositol diffused rapidly to the entire membrane, whereas PIP2 was confined to the membrane patch inside the pipette. The confinement of PIP2 was disrupted after cytochalasin D treatment, suggesting that the cytoskeleton is responsible for the low mobility of PIP2. The diffusion coefficient (D) of PIP2 in the plasma membrane measured with the fluorescence recovery after photobleaching technique was 0.00039 microm2/s (n = 6), which is markedly lower than D of phosphatidylinositol (5.8 microm2/s, n = 5). Simulation of PIP2 concentration profiles by the diffusion model confirms that when D is small, the kinetics of PIP2 depletion at different distances from phospholipase C becomes similar to the characteristic kinetics of GIRK inhibition by different agonists. These results imply that PIP2 depletion is localized adjacent to GqPCRs because of its low mobility, and that spatial proximity of GqPCR and the target protein underlies the receptor specificity of PIP2-mediated signaling.
Figures
References
-
- Hilgemann, D. W., Feng, S. & Nasuhoglu, C. (2001) Science STKE 111, RE19. - PubMed
-
- McLaughlin, S., Wang, J., Gambhir, A. & Murray, D. (2002) Annu. Rev. Biophys. Biomol. Struct. 31, 151-175. - PubMed
-
- Augustine, G. J., Santamaria, F. & Tanaka, K. (2003) Neuron 40, 331-346. - PubMed
-
- Nasuhoglu, C., Feng, S., Mao, Y., Shammat, I., Yamamato, M., Earnest, S., Lemmon, M. & Hilgemann, D. W. (2002) Am. J. Physiol. 283, C223-C234. - PubMed
-
- Irvine, R. (2004) Curr. Biol. 14, R308-R310. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
